

Did Natural Selection Play a Role in Speciation?
How Species Arose after the Ark
In our series, we’ve concluded that many new species have formed from the kinds that Noah took on board the Ark. We also observed that, when God created the kinds, He frontloaded them with genetic differences—with the potential to form all sorts of new species and varieties.
This fact alone completely transforms Charles Darwin’s “mystery of mysteries”—the origin of species.1 Darwin had no concept of our understanding of modern genetics, and the conclusions we have reached were entirely inaccessible to him. Specifically, with respect to how species change and the ultimate source of the visible varieties we observe, Darwin had no genetic insights. Conversely, since species are defined by their heritable traits, our modern genetic discoveries represent the first real, comprehensive answer to the central question Darwin pursued.
Our previous post left one significant detail unaddressed. While we carefully elucidated the origin of the vast majority of genetic varieties observable today, we didn’t quite connect the dots to how these varieties in individuals become distinct populations of new species.
From Genetic Differences to Visible Differences
In prior posts, we argued that, in individual members of the various kinds, God created the two copies of DNA different from one another. In technical terms, this is referred to as being heterozygous. For new visible traits to arise from this created heterozygosity, all that appears to be required is a shift from heterozygosity to homozygosity. The prefix homo- denotes “same,” and a movement towards homozygosity is a change from a DNA state in which the two copies differ toward a DNA state in which the two copies are more and more similar to each other.
To clarify, a shift from heterozygosity to homozygosity does not necessarily involve every chromosome. Though the image below depicts the most extreme examples (e.g., completely heterozygous and completely homozygous), a partially homozygous/partially heterozygous state can also exist. For example, if some of the chromosome pairs end up identical while other pairs remain different, this would represent a partially homozygous/partially heterozygous state. In fact, homozygosity need not even be at the whole chromosome level. Tiny chunks of a chromosome pair can be homozygous while other sections are heterozygous. Thus, a shift from heterozygosity to homozygosity can happen over a broad range of chromosome pairs and at multiple levels of chromosome organization.
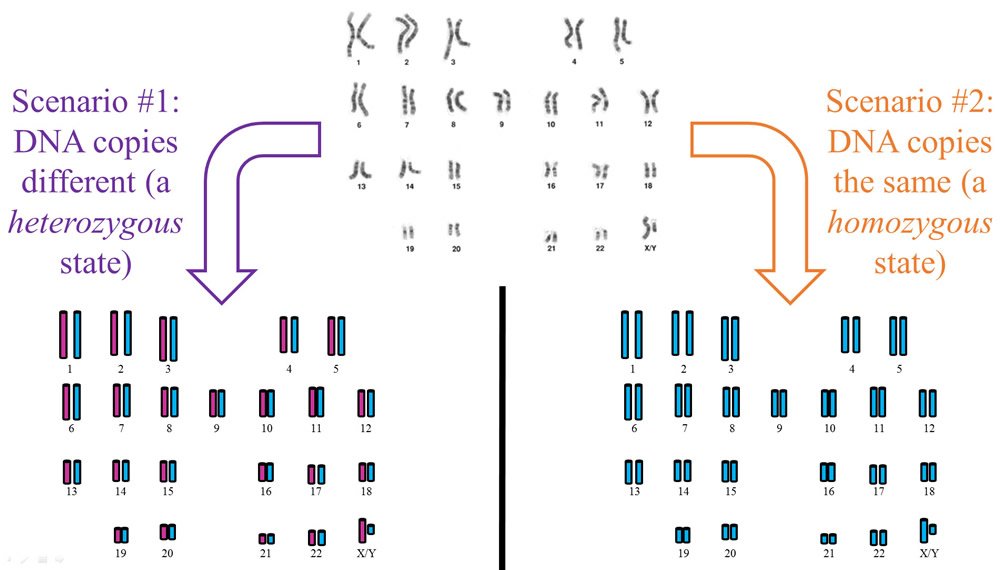
Figure 1. Human DNA is inherited from both parents. Hence, the condensed forms of DNA (visible in the top part of this image as floppy, noodle-like structures called chromosomes) come in pairs—one of each pair is inherited from each parent. Because our parents are different, each member of a chromosome pair is different from the other, as under scenario #1 on the left part of this image. Since Adam and Eve were created directly by God without human parents, it’s possible that both members of each chromosome pair were created identical, as in scenario #2 on the right. Nonetheless, available evidence suggests that each member of a chromosome pair was created different from the other. Under this scenario, it’s very easy to explain the millions of DNA differences that exist among humans today. Image of karyotype by Mats Halldin, via Wikimedia Commons.
Producing a more homozygous state is not hard. Gregor Mendel’s experiments with pea plants are instructive in this respect. At the visible level, Mendel observed that some traits can be hidden in one generation and appear in the next. For example, two pea plants that produce green, smooth-shaped pea pods can be crossed, and their offspring are a mix of 1) green and smooth-shaped pea pods, 2) green and rough-shaped pea pods, 3) yellow and smooth-shaped pea pods, and 4) yellow and rough-shaped pea pods. The information for yellow pea pods and rough-shaped pea pods was present in the parents, but it was masked. Green and smooth were dominant over yellow and rough.
Though Mendel was unaware, we now understand this phenomenon in more specific DNA terms. Since DNA is present in two copies (e.g., in chromosome pairs), even in pea plants, we can infer the DNA makeup of the green, smooth-shaped parental pea plants that gave rise to the same, as well as to yellow or rough-shaped pea pods. In short, for yellow or rough-shaped pea pods to appear consistently in the offspring of green and smooth-shaped pea pod parents, both parents must be heterozygous for the information for pea pod color and shape. In other words, in one pair of chromosomes, one copy of DNA in the parent must specify “yellow” and the other must specify “green.” Similarly, in another pair of chromosomes, one copy of DNA in the parent must specify “rough” and the other must specify “smooth.” When this heterozygous state exists, only green and smooth-shaped appear—they are dominant over yellow and rough-shaped.
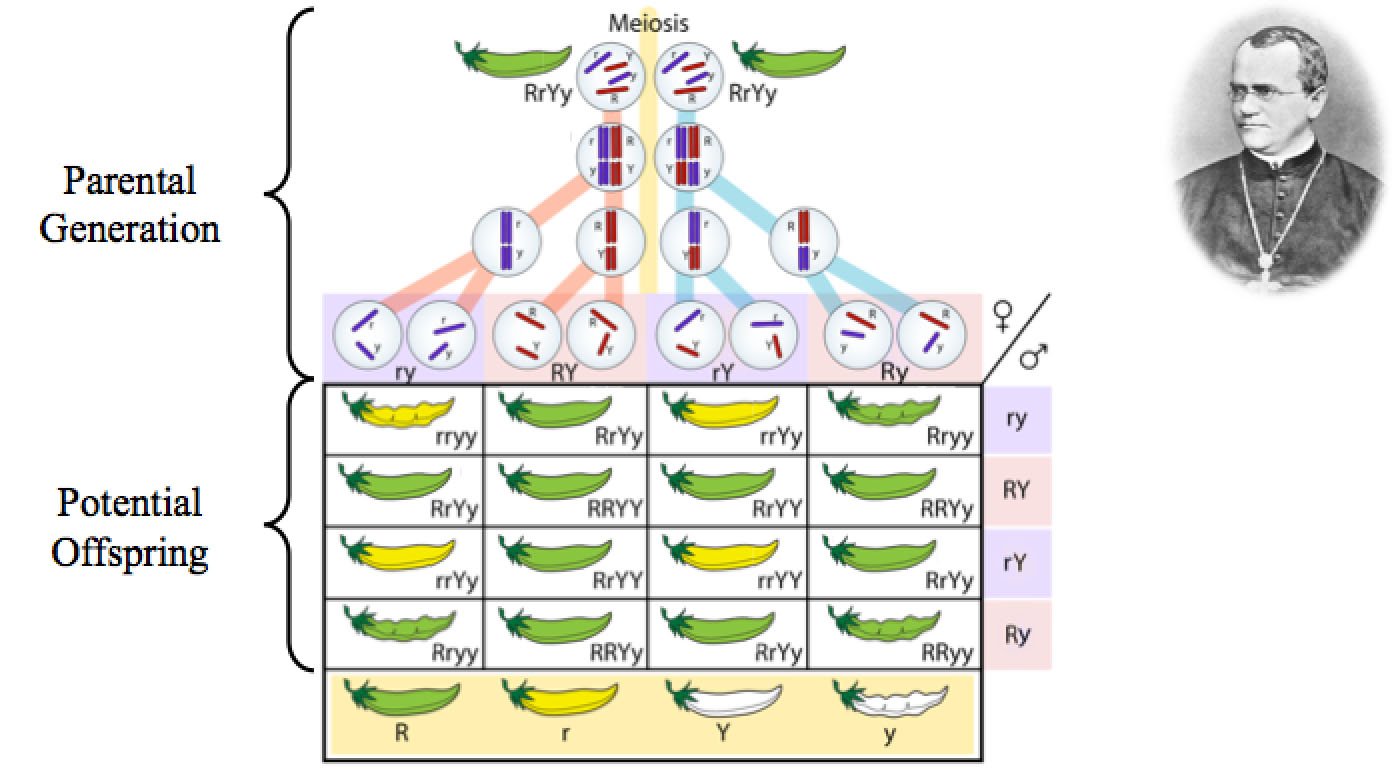
Figure 2. Gregor Mendel, the Austrian monk whose meticulous experiments in pea plants laid the foundation for our modern understanding of genetics, demonstrated that genetic potential can be hidden. Because DNA in the cell compartment (termed the nucleus) is inherited from both parents, it comes in two copies, which is illustrated by the two pairs of bars (representing chromosomes) that come in either a red or purple version. Information for pea pod color (R for green or r for yellow) and pea pod shape (Y for smooth and y for rough) is encoded by these bars, and each individual member of a bar pair need not encode the same information for shape and color—e.g., the top cells have all four versions (R, r, Y, and y). Nevertheless, despite having genetic information for yellow and rough pods, the appearance of the parents is green and smooth. During the process of cell division (termed meiosis), individual members of these chromosome pairs are distributed into sperm and egg cells, which permits a variety of combinations to arise in the offspring (e.g., green and rough-shaped pea pods, green and smooth-shaped pea pods, yellow and rough-shaped pea pods, and yellow and smooth-shaped pea pods). Image of Gregor Mendel by Materialscientist, via Wikimedia Commons; image of pea plant diagram adapted from LadyofHats, via Wikimedia Commons.
In the process of crossing the pea plants, only one copy of the each DNA pair is passed on from parents to offspring.2 If one or both of the copies carries the information for green pea pods, then offspring will have green pea pods. However, if the copy from one parent contains the instructions for yellow pea pods, and if the copy from the other parent also contains the instruction for yellow pea pods, the DNA information in the offspring will be only yellow. Since no information for green is present, the information for yellow is no longer hidden or masked by the dominance of the information for green pea pods. Hence, when two heterozygous parents are crossed, visible changes in traits can appear in a single generation.
For a new species to form, in technical terms, homozygosity must be maintained.
For this process to lead to the formation of a new species, a new population must be formed. If a new population does not form, the yellow offspring might cross with green offspring, and the next generation will have the yellow trait hidden again. Under this scenario, no permanent change will have taken place. For a new species to form, in technical terms, homozygosity must be maintained.
How could a homozygous state be isolated and kept from being mixed with heterozygous individuals? How could yellow pea pod offspring be prevented from crossing with green pea pod individuals?
In Gregor Mendel’s case, he personally oversaw the breeding process. Similarly, in animals, human breeders keep desired offspring from mating with other individuals that lack the desired traits.
But in the wild, no human breeders exist. How could homozygosity be maintained?
Unlike plants, animals can move or migrate. Homozygous animal individuals can easily be isolated from heterozygous individuals by migration. Not surprisingly, many species today are geographically isolated from one another.
For example, as their names imply, the African and Asian elephants exist on different continents. As another example, the 7 wild species of horse-like creatures that exist today—including three different species of zebras—are spread out over Africa and Asia.3 In the cat family, tigers are Asian, lions are primarily African, jaguars and pumas are American. In short, on a globe as big as ours, geographic isolation is easy.
Notice that we haven’t discussed survival of the fittest. For homozygous individuals to be isolated away from heterozygous individuals, the death of the original heterozygous individuals is not required. Migration of homozygous groups away from heterozygous groups would do the job just fine.
Naturally and by chance, some individuals will die, and others will survive to reproduce.4 As we observed in a previous post, the vast majority of mammal kinds died permanently—they’re extinct. But repeated cycles of massive population death followed by survival of a few individuals to found a new population are not necessary for speciation. Once God created kinds with enormous genetic variety from the start, reproduction and migration were virtually all that were needed to produce a huge number of species.
Speciation from Start to Finish
When God created the kinds heterozygous, He virtually guaranteed the formation of new species.
Let’s put the pieces together one more time to understand how easily species formed post-Flood. When God created the kinds heterozygous, He virtually guaranteed the formation of new species. The statistics of reproduction5 ensure the appearance of new traits in one or a few generations, and simple population growth curves indicate that these offspring can found new populations in short order. As these populations moved away from one another geographically, new species could form.
Is this process still occurring today? Take the African Cape buffalo as an example. Its curved horns have become a symbol of African wildlife. Yet several subspecies of the Cape buffalo exist. These subspecies also happen to be geographically distributed across the African continent. In perhaps a few decades, it wouldn’t be surprising if scientists labeled these subspecies as separate species.

Figure 3. Identity and geographic distribution of the three subspecies of the African Cape buffalo. Image of subspecies caffer by Nevit, via Wikimedia Commons; image of subspecies nanus by Llez, via Wikimedia Commons; image of subspecies brachyceros by Haster, via Wikimedia Commons; image of range map by Altaileopard, via Wikimedia Commons.
If this occurs, most people in the professional scientific community would probably view this as simply a bookkeeping change. Yet the process of speciation that I outlined above suggests that, in fact, what we’re observing right now is a bona fide formation of a new species.
Subspecies exist in many other species.6 Hence, speciation could be happening right now all over the planet. If we’re willing to consider the biblically consistent, scientifically justified model that I outlined above, I think we’d see a complete replacement for Darwin’s answer.
To underscore this fact, it should be clear from all that we’ve discussed that young-earth creationists are not evolutionists. We’re not a spin-off or an odd extension of Darwin’s principles. Instead, we postulate a very different source for the genetic variety we see today, and we explain speciation on a very different timescale.
Under the parameters we just laid out, evolution as Darwin described is not possible.
Furthermore, this front-loading of genetic information at the creation event also naturally sets limits on the speciation process.7 Since most of the genetic variety we see today goes back to the Creation Week, formation of new kinds (i.e., higher categorizations, such as at the Family level, not species) would require a massive—miraculous—input of new genetic information. Under the parameters we just laid out, evolution as Darwin described is not possible. In contrast, formation of new species from the kinds on board is not only possible, it represents a scientifically superior explanation to any that Darwin or his scientific descendants have proposed to date.
Ed. note: Figure 2, its caption, and referencing text have been slightly modified from the original version of this article that appeared on 7/30/16.
Footnotes
- Charles Darwin, On the Origin of Species, 1st edition, (London: John Murray, 1859), 1. http://darwin-online.org.uk/content/frameset?itemID=F373&viewtype=text&pageseq=1.
- Recombination of the DNA happens before the DNA copy is passed on, but this technical detail is not critical to our discussion at this point.
- Nathaniel Jeanson and Jason Lisle, “On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity: Genetic Clocks, Population Growth Curves, and Comparative Nuclear Genome Analyses Suggest Created Heterozygosity in Combination with Natural Processes as a Major Mechanism,” Answers Research Journal 9 (2016): 81–122, https://answersingenesis.org/natural-selection/speciation/on-the-origin-of-eukaryotic-species-genotypic-and-phenotypic-diversity/.
- Nathaniel Jeanson and Georgia Purdom, “Understanding Natural Selection: Clarifying the Confusion,” Answers in Genesis, February 16, 2016, https://answersingenesis.org/natural-selection/understanding-natural-selection/.
- E.g., as illustrated in Gregor Mendel’s crosses with pea plants.
- E.g., the common chimpanzee, the takin, the wildebeest, and so forth.
- Nathaniel Jeanson and Jason Lisle, “On the Origin.”

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis