

Getting Enough Genetic Diversity
How Species Arose After the Ark
In this series, we have derived the fact of speciation within the young-earth creation view, and we have observed several lines of evidence supporting or consistent with the recent origin of species.
The major remaining question is how tens of thousands of species could arise in a few thousand years.
Recasting a Common Objection
To skeptics of the Scriptures, producing so many species in such a short time span—short as compared to the evolutionary time span—seems implausible. Laying aside both Darwin’s breed-species analogy and the mitochondrial DNA clock data, critics of young-earth creation don’t see how biological change can happen so quickly. To them, we haven’t witnessed 10 species form in the last few thousand years, let alone tens of thousands.
These skeptics forget the evidence right under their nose. While the origin of over 30 living cat species might appear to require significant morphological changes, visible variety of a much greater magnitude can arise in an even shorter frame of time. For example, a frog begins its life as a fertilized egg—as single cell. No limbs, head, eyes, tongue, long toes, and so on are visible at the single-cell stage. Yet, in the process of development and maturity that spans less than 3 years, these anatomical features are formed. This progression from a single cell to an adult frog represents far more visible change than the process of forming new species from the kinds on board the Ark. Thus, when the skeptic claims that we’ve never witnessed the amount of morphological change required by the young-earth speciation model, the skeptic does so by ignoring the familiar process of development.
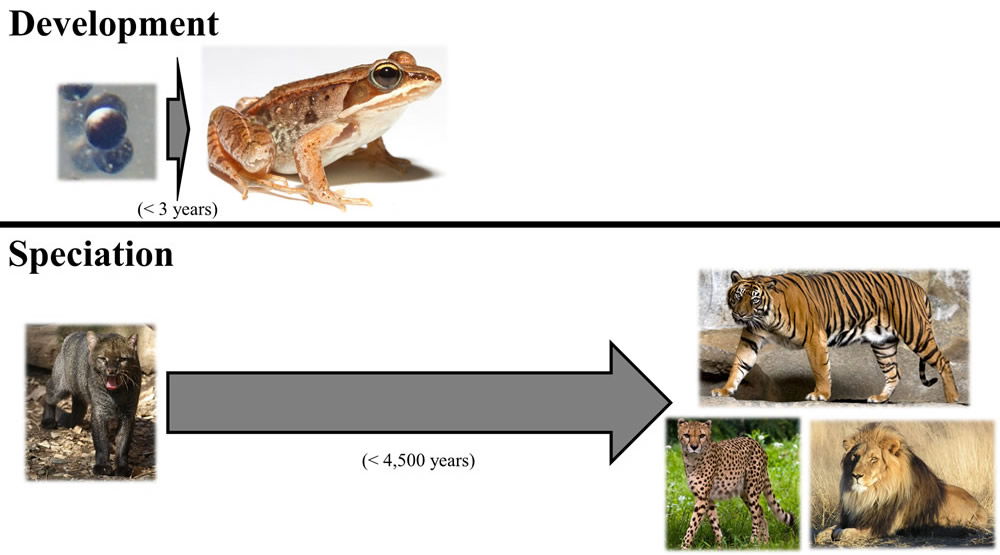
Image adapted from figure 2 of Nathaniel Jeanson and Jason Lisle, “On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity: Genetic Clocks, Population Growth Curves, and Comparative Nuclear Genome Analyses Suggest Created Heterozygosity in Combination with Natural Processes as a Major Mechanism,” Answers Research Journal 9 (2016): 81–122, https://answersingenesis.org/natural-selection/speciation/on-the-origin-of-eukaryotic-species-genotypic-and-phenotypic-diversity/.
Nevertheless, the skeptic might insist that the process of development is primarily controlled by a different set of instructions than the process of speciation. This claim is indeed true. Development proceeds according to the epigenetic information in the cell,1 but speciation is a primarily genetic phenomenon. Therefore, when discussing speciation post-Flood, our primary concern should not be the formation of new visible features; it should be the origin of genetic differences among species.
The Genetics of Species’ Origins
Can enough genetic diversity arise in a few thousand years to make the young-earth speciation model work? Today, millions of DNA differences separate species from one another.2 How can millions of DNA differences arise in a few thousand years? To the skeptic, the answer is simple—millions of DNA differences can’t.
However, this claim is ill-informed. One of the genetic observations that we made previously is all the more relevant here. In a prior post, we observed that mitochondrial DNA acts like a clock. As the rungs of the DNA ladder change with time, the length of time that a species has been on the planet is recorded.
When we perform a similar clock analysis on the rest of the DNA sequence (e.g., the sequence in a cellular compartment termed the nucleus), we don’t reach the same conclusion. For example, using the rate at which copying errors are made in the billions of DNA letters in the human nuclear DNA sequence, it is not possible to produce millions of DNA differences in a few thousand years.3 Similar results hold true in other species.4
Image by MesserWoland, via Wikimedia Commons.
At first pass, this may suggest that one of the clocks must be wrong. Upon further reflection, we reach a different conclusion.
Consider a simple kitchen example as an analogy. Let’s say that, one afternoon, you happen to glance at the oven clock, and the time shows 2:57. Let’s also say that, in the same moment, you happen to glance at the display on the microwave oven, and it shows 1:22. At first pass, these might appear to contradict.
With a little more information and contemplation, it’s not hard to see how both of these timing devices display accurate information. The oven clock measures how much time has passed since noon; 2 hours and 57 minutes have elapsed. In contrast, the time display on the microwave represents the remaining minutes and seconds on the timer—1 more minute and 22 seconds will expire before the timer sounds. Both display a correct measure of time, but each begins from a different starting point and has a different goal.
In a similar way, the mitochondrial DNA and the nuclear DNA sequences both measure time, but they do so from different starting points. Unlike most of the DNA in our cells, mitochondrial DNA is inherited primarily—if not exclusively—through the maternal line. Though males and females both possess and require mitochondrial DNA, only moms pass on mitochondrial DNA to their offspring. In contrast, the DNA in the nucleus comes from both parents.
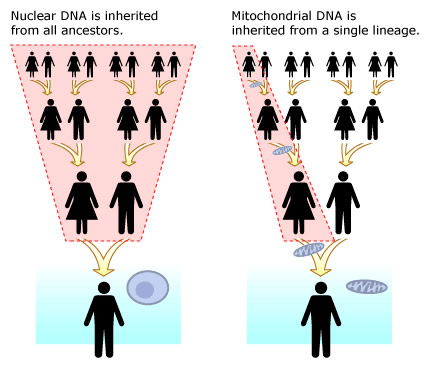
Image by PaleoBioJackie, via Wikimedia Commons.
This latter biological fact illuminates another potential source of DNA variety. When God created Adam and Eve, He would have created them both with mitochondrial DNA. However, only Eve would have passed on her mitochondrial DNA to her offspring. All the differences we see today are the results of errors in copying Eve’s original sequence.5
When God created the nuclear DNA in Adam and Eve, He created it in two copies as 23 paired chromosomes. Even though Adam and Eve did not have human parents from which to inherit two copies, it is likely that God created their two nuclear DNA sequences different from one another. In other words, Adam’s first set of 3 billion DNA letters would have contained a certain sequence, and his second set of 3 billion DNA letters would have contained a different sequence—with perhaps 5–10 million differences between the two sets.6 This would allow them to be fruitful, multiply, and produce diverse offspring—instead of producing clones.
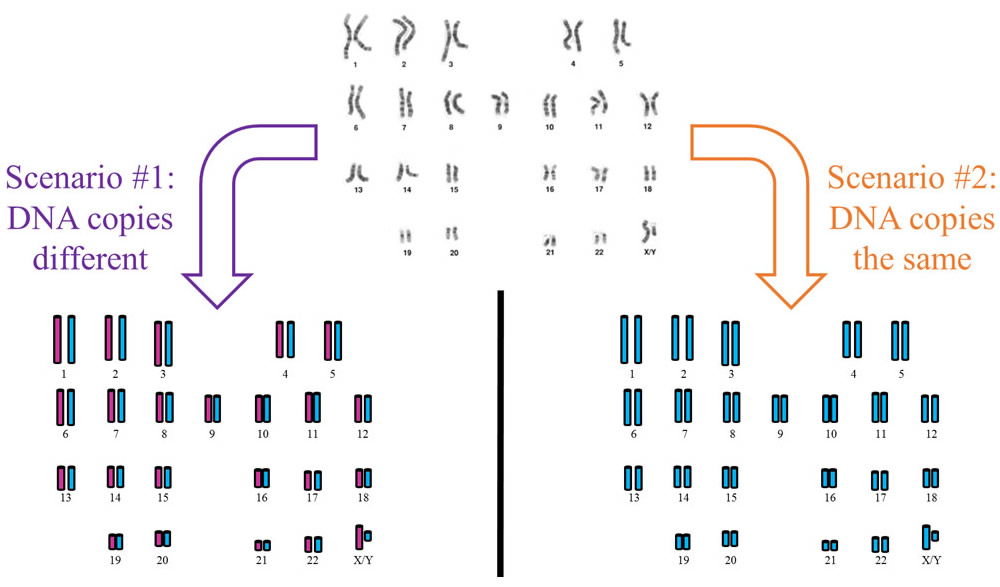
Top center image by National Human Genome Research Institute, via Wikimedia Commons.
What I’ve just described also makes testable scientific predictions, which just happen to fit what we observe today when we compare modern human nuclear DNA sequences.7
Thus, the two DNA clocks don’t contradict. Mitochondrial DNA comparisons don’t contradict nuclear DNA comparisons. Both clocks tell the same time once we carefully account for their differing starting points.
Conversely, the nuclear DNA starting point I just described easily accounts for the millions of DNA differences that exist among humans today. Copying errors are unable to explain the vast amount of DNA differences we see today because the vast majority of these differences are not the product of error, but of deliberate design.8
Similarly, it appears that God created nuclear DNA differences in the animal kinds as well. Genetic calculations suggest that God created tens of millions of DNA differences from the start. In other words, the cat kind ancestor was likely created with one DNA copy different from the other DNA copy—with, perhaps, 50 million DNA differences between the two copies.
Since only about 1,660 years passed between the Creation event and the Flood, most of this variety would have been carried on board the Ark. Being so shortly removed from the perfect creation, it’s not hard to believe that significant population growth in these animal kinds occurred. As we observed in a previous article, massive amounts of population growth can happen in a very short amount of time, even in slowly reproducing species. Under this scenario, straightforward population modeling indicates that very little of the created genetic differences would have been lost prior to the Flood.9
Implications of Created DNA Differences
Once the animals stepped off the Ark, their reservoir of DNA differences could have easily translated into a massive amount of morphological change. This fact is best illustrated by a consideration of a recent objection to the young-earth creation view of speciation. Over a decade ago, one of the main complaints that could be levied was based on the function of the DNA letters. Actually, it was the apparent lack of function of DNA differences. When the sequence of the billions of human DNA letters were first elucidated in 2001, many in the scientific community concluded that the vast majority was “junk”—useless leftovers from the human evolutionary process. Presumably, similar amounts of “junk” DNA existed in other species as well.
However, this claim was not based on rigorous laboratory testing of the billions of DNA letters in the human DNA code. Instead, it was based on appearance—to evolutionary scientists, the DNA “looked” as if it were non-functional. Since 2001, a large body of laboratory (e.g., experimental) evidence has accumulated. While preliminary and largely biochemical, this evidence has a trajectory suggesting that that vast majority of human DNA is, in fact, functional. With a few more years of experiments, the evidence for function will likely increase.
In addition, this trajectory seems to be true of other species. As more functional testing has been performed in non-human species, more evidence for DNA function has accumulated. Hence, the long-held ideas of how DNA works and how much of it is functional have undergone a massive shift within the last decade.10
These results have profound ramifications for the speciation question. Since DNA now appears to be extremely functional, it would seem that the production of a new species would simply require a few DNA differences to arise.
In fact, as we observed earlier, millions of DNA differences separate species within a kind.11 A massive potential for speciation exists all over this planet.
A Comprehensive Model
Let’s put all the pieces of this genetic puzzle together. Mitochondrial DNA is inherited uniparentally—from only one of the two parents, the mother. Consequently, it is a very effective yet simple time keeper for elucidating the time when species originated. As copying errors have occurred over multiple generations, the mitochondrial DNA “clock” has ticked off the time since species first arose. This clock indicates that species originated within the last few thousand years.
The rest of the DNA sequence within a creature (e.g., the nuclear DNA) is inherited from both parents. Because of the biparental manner (e.g., from both parents) in which this DNA is inherited, God could have created the kinds with DNA differences from the start, and at least two independent lines of evidence argue that this is true.12 Thus, nuclear DNA also records a timeline pointing to a recent origin for species, a timeline that makes testable predictions.
Conversely, the fact that so many nuclear DNA differences were present from the start has tremendous implications for the plausibility of speciation on the young-earth timescale. With millions of DNA differences—massive amounts of DNA variety—encoded into each kind from the start of their existence, the potential for speciation is mind-boggling. Combined with the fact that species can recover enormous population sizes in very short amounts of time, in addition to the fact that the vast majority of DNA sequences within a creature appear to be functional, these results demonstrate that millions of species in a few thousand years is not only plausible, it is also probable.



Footnotes
- Epigenetics is a scientific field concerned with heritable information that exists in addition to the information encoded by the linear sequence of DNA letters.
- Nathaniel Jeanson and Jason Lisle, “On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity: Genetic Clocks, Population Growth Curves, and Comparative Nuclear Genome Analyses Suggest Created Heterozygosity in Combination with Natural Processes as a Major Mechanism,” Answers Research Journal 9 (2016): 81–122, https://answersingenesis.org/natural-selection/speciation/on-the-origin-of-eukaryotic-species-genotypic-and-phenotypic-diversity/.
- This assumes that the rate at which copying errors are made has been constant.
- Jeanson and Lisle, “On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity.”
- Copying errors would compound with time. One of Eve’s descendants would have made a copying error, and her offspring would have used this erroneous template from which to copy again—and likely introduce even more copying errors. See Nathaniel T. Jeanson, “On the Origin of Human Mitochondrial DNA Differences, New Generation Time Data Both Suggest a Unified Young-Earth Creation Model and Challenge the Evolutionary Out-of-Africa Model,” Answers Research Journal 9 (2016):123–130, https://answersingenesis.org/genetics/mitochondrial-dna/origin-human-mitochondrial-dna-differences-new-generation-time-data-both-suggest-unified-young-earth/.
- Since Eve was made from Adam’s side, she might have had DNA identical to Adam, except for the Y (male) chromosome. In other words, her two copies of DNA would have been different from one another—but nearly identical (except for the Y chromosome) to Adam’s two copies of DNA. Eve’s sex chromosome pair would have been XX, while Adam’s sex chromosome pair would have been XY. In the remaining 22 pairs of chromosomes, DNA differences would have existed within Adam and Eve, but hardly any DNA differences (except for the male- and female-specific DNA differences) would have existed between Adam and Eve. This perspective is slightly different from one that was suggested previously, but it seems to be the best fit to our current understanding of global human DNA diversity (see Robert W. Carter, “The Non-Mythical Adam and Eve!,” Creation Ministries International, August 20, 2011, http://creation.com/historical-adam-biologos).
- Jeanson and Lisle, “On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity.”
- Copying errors since the creation of Adam and Eve’s DNA sequences would also contribute to the DNA differences that exists today, but it would represent a small fraction of the total amount of DNA differences. See Jeanson and Lisle, “On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity.”
- Carter, “The Non-Mythical Adam and Eve!” See also Robert W. Carter and Matthew Powell, “The Genetic Effects of the Population Bottleneck Associated with the Genesis Flood,” Journal of Creation 30 (2016): 102–111.
- Jeanson and Lisle, “On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity.”
- Ibid.
- Due to the genetic complexity of the second line of evidence, I omitted it from the present lay-level summary article. It can be found in Jeanson and Lisle, “On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity.”

{kind=link}
{kind=link}
{kind=link}
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis