

Chapter 27
Is Speciation Evidence for Creation or Evolution?
by
Dr. Gary Parker
on
October 3, 2020
Republished with permission and featured in
The New Answers Book 4
In a debate at a major Texas university, the creationist was challenged with this claim: Hawaiian fruit flies that could once all interbreed had changed into numerous reproductively isolated species, and that, said the challenger to considerable applause, “proved evolution.” The creationist responded (also to considerable applause) that such a change would be the opposite of evolution. Losing the ability to interbreed, each “new species” would have less genetic variability, less ability to meet changes in its existing environment, and less ability to explore new environments — all suggesting decline and demise rather than the expansion of genetic potential required for what Darwin called “the production of higher animals.”
Which of these views is more consistent with our present understanding of genetic science and with the biblical record of earth history?
Basic Genetics
The Bible records several key events in early earth history that suggest concepts geneticists can test scientifically. Genesis 1 states that God created many distinct “kinds.” We infer from a plain reading of Scripture that animals and plants were created to reproduce within the boundaries of their kinds (Genesis 1, 6, and 8). A created kind is typically equivalent to the level of family in modern classification schemes as many members of a family can interbreed and produce offspring. The kinds were also “to fill” (scatter, move into) earth’s varied environments (Genesis 1:22, 8:17). Multiple biological mechanisms accounted for this filling and resulted in variation within kinds, or speciation. Do these fundamental concepts in God’s Word — discrete created kinds, or baramins (Hebrew: bara = create and min = kind), having broad but limited variability — help scientists understand the genetic changes in organisms and speciation found in God’s world? Indeed, they do!
The complete set of DNA specifying a kind is called its genome. The human genome includes approximately 20,000 to 25,000 protein-coding chromosomal segments commonly called genes. The genes and the information they encode are largely responsible for the set of biological traits that distinguish human beings from other kinds of life. All humans have essentially the same genes, and they are over 99 percent similar in all seven billion of us; hence, geneticists refer to the human genome and have concluded that we are all members of one race, the human race (as the Apostle Paul preached in ancient Greece, Acts 17:26).
The similarity among all human beings is obvious, but so is the tremendous variation!
The similarity among all human beings is obvious, but so is the tremendous variation! The genes we share in the human genome make us all the same (100 percent human), but different versions of these shared genes, called alleles, produce the spectacular variation that makes each individual unique. For any given gene, God could have created it in four different allelic varieties (two in both Adam and Eve). Genetic alterations occurring since sin corrupted creation have introduced many new alleles, but no new genes.
The human genome, for example, has genes for producing hair and controlling its shape; allelic versions of these genes result in individuals with straight, wavy, curly, and tightly-curled hair; all variations within the human kind. Although, genetically speaking, skin color is more complex, the variation in human skin tone can be described as the action of two pairs of genes with different alleles (A/a and B/b) that influence the production of the skin pigment melanin. As shown in figure 1,1 two people with medium-brown skin tone and genes AaBb could have children with the full range of skin tones — from very dark (AABB), to dark (AABb or AaBB), to medium (like AaBb), to light (like Aabb), to very light (aabb). That would certainly be “change through time” but a lot of change in a little time (one generation!) With no genes added, this is just variation within a kind.
Mutations, changes in DNA that occurred after man’s sin corrupted God’s creation, do not produce new genes. Rather, mutations only produce alleles, variations in pre-existing genes. Alleles are not different genes in the sense that genes for skin color and genes for making sickle-cell hemoglobin (resulting in sickle-cell anemia) are. Similarly, the sickle-cell gene is a different allele (version) of the hemoglobin gene in the sense that it was not present at creation, but it is only a different harmful version of a pre-existing gene. In fact, the allele for sickle-cell hemoglobin differs in sequence in only one position out of several hundred from the normal gene for making hemoglobin. Again, we see mutations leading to different versions of pre-existing genes resulting in a variety of alleles but not the creation of brand new genes encoding novel proteins with novel functions of the type necessary for molecules-to-man evolution.
Variation Within a Kind
All the genes in one generation available to be passed on to the next are called the gene pool. Members of the same kind may also be defined as organisms that share the same gene pool. The number of genes for different kinds of traits, the number in a complete genome, can be called the depth of the gene pool. The human gene pool is around 20,000–25,000 genes deep. The width of the gene pool refers to the amount of its allelic variation. Among dogs, for example, the width of a greyhound’s gene pool is very narrow; crossing purebred greyhounds just gives you more greyhounds, all very similar in speed, color, intelligence, hair length, nose length, etc. Crossing two mongrels, however, can give you big dogs and small dogs, dark and light and splotchy-colored dogs, dogs with long and short hair, yappy and quiet dogs, mean and affectionate dogs, and the list goes on! The width of the mongrel’s gene pool (its allelic variability) is quite large compared to the greyhound’s, but the depth of the gene pool (the number of genes per genome) is the same for both dogs.
A kind is defined in terms of depth of the gene pool, which is the total number of different genes in a genome and a list of traits they encode for. Variation within a kind is defined in terms of the width of the gene pool, the number of possible alleles at each gene site (locus).
Geneticists call the shuffling of pre-existing genes recombination. Perhaps you have played a game with a common deck of 52 cards that includes four groups (hearts, diamonds, clubs, and spades), each with 13 different numbers or “faces” (2–10 plus J, Q, K, A). In a game called bridge, each of four players gets a “hand” of 13 cards. You can play bridge for 50 years (and some people do!) without ever getting the same group of 13 cards. The hands you are dealt are constantly changing, and each is unique — but the deck of cards remains always the same.
Although the comparison is not perfect, a deck of cards illustrates the concept of variation within a created kind. The bridge hands dealt are unique, different, and constantly changing, like the individual members of a population. But the deck of 52 cards remains constant, never changing, always the same, like the kind. Individual variation plus group constancy equals variation within a created kind.
Faith in Man Versus Faith in God’s Word
Based on faith in Darwin’s words, evolutionists assume that all life started from one or a few chemically evolved life forms with an extremely small gene pool. For evolutionists, enlargement of the gene pool by Darwinian selection (struggle and death) among random mutations is a slow, tedious, grim process that burdens each type with a staggering “death load” and “genetic load” of harmful mutations and evolutionary leftovers. Based on faith in God’s Word, creationists assume each created kind began with a large gene pool, designed to multiply and fill the earth with its tremendous ecological and geographic variety.
Neither creationists nor evolutionists were there at the beginning to see how it was done, of course, but the creationist can build on the Word of the One who was there “in the beginning” (Genesis 1:1; John 1:1–3). Furthermore, the creationist mechanism is consistent with scientific observation. The evolutionary mechanism doesn’t work and is not consistent with present scientific knowledge of genetics and reproduction. As a scientist, I prefer ideas that do work and do help to explain what we can observe, and that’s biblical creation!
Since animals were commanded to multiply and fill the earth, we can infer that the created kinds were “endowed by their Creator” with tremendous allelic variability and allelic potential in very wide gene pools. Geneticists now know, for example, that alleles for the full range of normal human variation — darkest to lightest skin tone, Pygmy to Watusi heights, wide to thin lips, hair from straight to wavy to curly to tightly-curled, eyelids producing round to oval shapes, etc. — are possible, beginning with just two people. Genetics problems solved by high school students (figure 1) show how such parents could produce children with traits from darkest to lightest, shortest to tallest, with hair of any style, and eyes and lips of any shape in just one generation — all with NONE of the deep time, chance mutations, and ceaseless struggle to the death that evolutionists use to explain variation in beak sizes in finches or amounts of black pigment in moth wings.
What Does This Awesome Variability Within Kinds Mean?
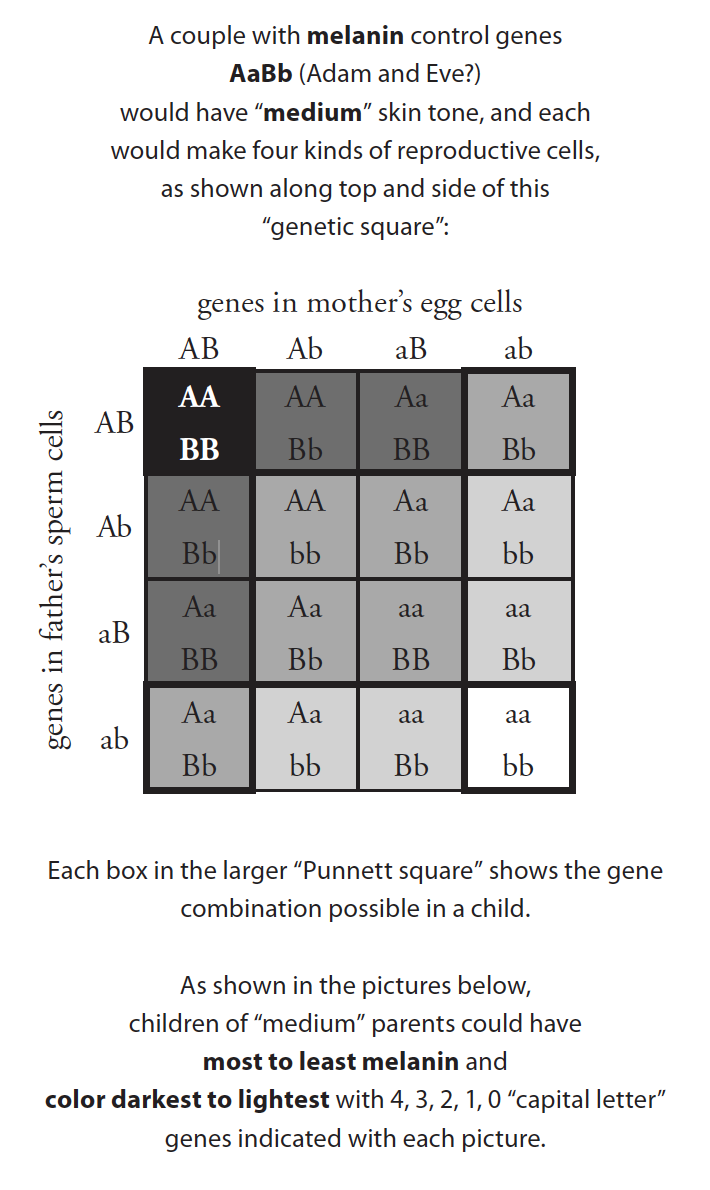
Figure 1. Inheritance of melanin skin color
We still see God’s creativity unfolding before our very eyes in a different way in the birth of each child. As they relate to the genetic potential God created in our first parents, we may not yet have seen the fastest runner or the greatest mathematical or musical genius.
For one thing, such awesome variation reflects God’s creativity. God created the first man from the dust of the ground and the first woman from a rib from his side (Genesis 2:7, 21–22). Then, God rested from His creative acts at the end of the creation week (Genesis 2:1–2). But we still see God’s creativity unfolding before our very eyes in a different way in the birth of each child. As they relate to the genetic potential God created in our first parents, we may not yet have seen the fastest runner or the greatest mathematical or musical genius. Genes were not produced one at a time by evolutionary processes — time, chance, mutations, struggle, and death over millions of years. This unfolding of genetic variability in pre-existing genes is all stunning variation within a kind, but it is NOT the formation of new genetic information of the type required for molecules-to-man evolution.
As the descendants of each created kind multiplied to fill the earth, we see their genetic potential unfolding. God created the bear kind, for example. But as bears moved into different environments around the world after the Flood, their built-in variability and ability to genetically change came to visible expression in black bears, brown bears, grizzly bears, polar bears, etc. The created dog kind diversified into specialized subtypes: wolves, coyotes, domestic dogs, etc. Think also about the tremendous genetic variability brought to visible expression in the cat kind, rose kind, tomato kind, etc.
There is a strong tendency, both in nature and in experimental breeding, for generalized, adaptable organisms to produce a variety of specialized, adaptable subgroups. Figure 1, discussed earlier, showed that if Adam and Eve, for example, had a variety of alleles for skin tone (AaBb) they could have children with skin tones from darkest to lightest. However, some of that initial genetic variability would be lost when subgroups of the human population moved apart and remained reproductively isolated, as they did at the Tower of Babel (Genesis 11). Some language groups may have included only A and B alleles, losing a and b; in such AABB subgroups, parents could only have children with very dark skin. Subgroups without the A and B alleles (only a and b) would produce only very light-skinned children, and either AAbb or aaBB subgroups would always be medium brown. AaBb subgroups would continue to produce the entire color range, like some groups in India still do today.
Darwin thought otherwise, but scientists now recognize that people groups who express only part of the full range of melanin color variation (such as very dark skin) are 100 percent human. But among animals and plants, both in nature and from selective breeding, subgroups of some kinds may become so different (e.g., size, courtship ritual, mating season, chromosomal rearrangements, aggressiveness, etc.) that they can no longer interbreed (even though their identity as members of the same created kind can still be confirmed by genetic testing). Such reproductive isolation was once used as the key criterion for defining species.
What? Two or more specialized species descended from one generalized ancestral kind? Doesn’t that prove evolution after all? Exactly the opposite.
Speciation, yes; evolution, no. Molecules-to-man evolution requires a net increase in novel genetic information, the addition of genes for new trait categories to a genome. Reproductive isolation and subsequent speciation result in a loss of genetic variability (alleles), converting a large gene pool into subgroups with smaller gene pools (i.e., “new species” with less ability to meet changes in their environment, restricted ability to explore new environments, and reduced prospects for long-term survival). Indeed, evolutionists now regularly use the term overspecialization in speciation as an explanation for extinction versus evolutionary progress.
The Florida panther, for example, is considered an endangered species. What endangers it? The small, inbred population was so riddled with mutations that no cubs could survive to reproductive age. The cure? Since it is only a species within a kind, it was bred with western panthers (members of the same kind) having different post-Fall mutations. The former Florida panther is now recovering from its flirt with extinction and being restored to health.
Distinctive genetic diseases and abnormalities characterize many purebred dogs, which have often reached the end of the line, genetically speaking. Each has all the genetic information in its genome to be 100 percent dog (so each has the same gene pool depth), but the allelic variability (gene pool width) could be reduced ultimately to 0 percent (only one allele per locus in a population). Therefore, crossing purebred poodles with poodles, for example, would produce only poodles and would not be a promising path for recapturing the ancestral wolf or generalized dog kind. If a “poodle plague” wiped out the poodle, however, poodles could be brought back again over several generations through breeding wolves or mongrel dogs. Even the quagga, an extinct subspecies of zebra, is being brought back through cross-breeding varied members of the horse kind.
The Wrong Kind of Change
Speciation is moving in the wrong direction to support the evolutionary belief in upward changes between kinds, or molecules-to-man evolution. Speciation produces only variation within kinds as a result of the subdivision and/ or alteration of pre-existing genetic variability. Speciation also brings to visible expression the magnificent variability and potential for variation that God programmed into the members of each of the original created kinds.
After man’s sin, mutations introduced many “negative variations,” helping scientists to explain the origin of birth defects and disease. Evolutionists had hoped mutations would provide the new genetic information required to move organisms up the so-called evolutionary tree. But mutations only produce variation in pre-existing genes, which are alleles that only make a gene pool wider rather than deeper. So mutations result in variation within a kind and not the formation of new and different kinds, which Darwin called the “production of higher animals.”
Uncritical acceptance of evolution has so stunted scientific thinking that people give mutations god-like qualities. They act as if a cosmic ray striking a cell can cause a mutation that somehow assembles over 1,500 DNA nucleotides into a brand new gene, regulators and all, that suddenly begins producing a brand-new protein responsible for a brand-new trait, raising the lucky mutated organism to the next higher limb on the evolutionary tree! NOTHING remotely like that has ever been observed, nor will it be!
Mutations have no ability to compose genetic sentences, no ability to produce novel genetic information, and, hence, no ability to make evolution happen, at all.
Mutations are NOT genetic “scriptwriters”; they are merely typographic alterations in a genetic script that has already been written. Typically, a mutation changes only one letter in a genetic sentence averaging 1,500 letters long. To make evolution happen—or even to make evolution a theory fit for scientific discussion—evolutionists desperately need some kind of genetic scriptwriter to create novel genetic information, increasing the size of a genome and the depth of a gene pool. Mutations have no ability to compose genetic sentences, no ability to produce novel genetic information, and, hence, no ability to make evolution happen, at all.
Yet molecules-to-man evolution requires phenomenal expansion of genetic information. It would take thousands of mutations adding novel information to change simple cells into invertebrates, vertebrates, and mankind. The evolutionist’s problem is with the fundamental nature of information itself. The information in a book, for example, cannot be reduced to nor derived from the properties of the ink and paper used to write it. Similarly, the information in the genetic code cannot be reduced to nor derived from the properties of matter or the allelic variations caused by mutations. Its message and meaning originated instead in the mind of its Maker, Jesus Christ, the Author of life (John 1:1–3). What we see in God’s world agrees with what we read in God’s Word.
The New Answers Book 4

Building on the previous New Answers Books, learn more about the Gospel and a young earth, death of plants and leaves, dragons, religious wars, cavemen, science, living fossils, and more.
Read Online Buy Book
Master Books has graciously granted AiG permission to publish selected chapters of this book online. To purchase a copy please visit our online store.
Footnotes
- Gary Parker, Building Blocks of Life Science (Green Forest, AR: Master Books, 2011), 9.
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis