

Tiktaalik and the Fishy Story of Walking Fish, Part 2
An in-depth look
Abstract
Now that the euphoria in the media has settled down on Tiktaalik, the “walking fish,” it is time to reflect upon what must surely be the biggest fish story of the decade.
Keywords: Tiktaalik, walking fish, evolution, air-breathing, ancestors, tetrapods, missing link, ichthys, Darwin fish, legs, Osteicthyii, bony fish, Sarcopterygii, lobe-fin fish, Dipnoi, lungfish, Crossopterygians, coelacanth, living fossils, Eusthenopteron, Panderichthys
Read part one in this series.
In both the print and broadcast media last year, reports of the discovery of the fossil fish known as Tiktaalik were hyped as convincing proof that, through a random chance process of evolution, fish sprouted legs and walked out onto the land where they turned into amphibians, reptiles, mammals, and, ultimately people. But the media’s excitement seems to stem not so much from being able to report a real scientific discovery as in being able to discredit the biblical account of Creation.
A front page article in The New York Times,1 for example, hailed Tiktaalik “as a powerful rebuttal to religious creationists, who hold a literal biblical view on the origins and development of life.”
The walking fish has become symbolic of evolutionism

The whole idea of walking fish has come to be symbolic of the evolutionary worldview and its opposition to biblical Christianity. Many evolutionists display the familiar “Darwin fish” symbol on their automobiles, T-shirts, and office doors as a public declaration of their allegiance to evolution. The “Darwin fish” is a desecration of the fish symbol used by early Christians as a means of mutual identification during a time of persecution. Christians chose the fish symbol because the individual letters of the Greek word ichthys (for “fish”) served as an anagram for “Jesus Christ Son of God, Savior.” Evolutionists have substituted the word “Darwin” for “ichthys” and have placed walking legs with feet on the fish. Thus, the Darwin fish reflects the fact that many evolutionists have indeed replaced Christianity with Darwinism. As for the legs on the Darwin fish, we will see that there are no known fish with true “legs” (and certainly no feet), and none capable of actually “walking”—except in the most trivial sense of the word.
We must be cautious of evolutionary claims
In the next months and years, there will doubtless be further claims in the popular media of “irrefutable proofs” for evolution and, more importantly, “proofs” against the biblical account of Creation. The popular media—as with tax-supported zoos, science museums, and public schools—are often zealous supporters of the quasi-scientific religion of materialism.
However, few reporters, teachers, or laymen have ever read the original scientific reports upon which grandiose evolutionary claims are based. Moreover, these reports are often convoluted, conflicting, and couched in unprovable assumptions that make evolutionary claims difficult to evaluate even for those who do examine the original scientific papers.
To evaluate the claims that there are fossil fish with legs that walked out of water to take up permanent residence on the land, one needs to understand something about fish, tetrapods (limbed vertebrates including humans), legs, and what is required anatomically to walk and swim. So, let us begin by looking at the wide world of fish, and see which ones are supposed to be the “walkers.”
There are lots of fish!
The first thing to consider is that there are a lot of fish—both living and fossilized. Approximately 25,000 species of currently living fish have been identified, with 200–300 new species being discovered—not evolved—every year. Indeed, fish comprise fully half of all known vertebrates!
Losing thousands of species of fish is hardly evolution—it’s extinction.
It is not clear how many different fish species have been found as fossils, but some experts claim that there were once nearly a million species of fish! It appears that over time we have lost a lot of species of fish—and retained relatively less.
But losing thousands of species of fish is hardly evolution—it’s extinction. The question is, have we really gained any fundamentally new fish (to say nothing of fish that evolved true legs and walked out onto the land as permanent residents)?
Classification of fish
Fish come in a bewildering variety of forms that defy consistent classification. As a result, there are competing classification schemes based on the particular bias of the classifier.
Basically, all species of fish have been divided into two main types—the jawless fish (hagfish and lampreys) and the jawed fish (all the rest). The jawed fish are in turn, divided into two groups: the cartilaginous fish (such as the sharks and rays which have a skeleton made of flexible cartilage) and the much more numerous bony fish, which have hard bony skeletons.
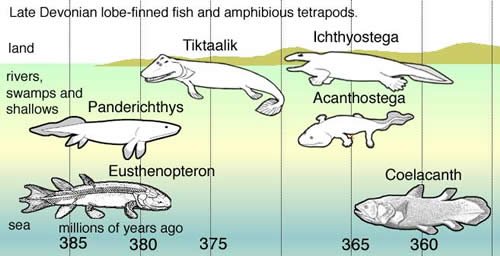
Wikimedia Commons
The evolutionary timeline of lobe-finned fish and amphibious tetrapods. Many of the so-called transitional forms have been greatly disputed, discovered (e.g. coelacanth), or dismissed, and Tiktaalik has recently been propped up as the “savior” of the evolutionary paradigm. How soon will it be before Tiktaalik is abandoned also?
Evolutionists believe that it took about 100 million years for invertebrates (animals with no bones) to evolve into vertebrates (animals with backbones). However, no compelling fossil evidence documents this purported major and unambiguous transition.
While evolutionists believe that fish were the first true vertebrates, they’re not sure which evolved first—cartilaginous or bony fish.
During the embryological development of vertebrates, most bones develop first as cartilage models that are later replaced by bone (called endochondral bone). Following the dictates of the embryonic recapitulation myth, it would be attractive for evolutionists to propose that cartilaginous fish evolved into bony fish, but most evolutionists consider the cartilaginous fish to be far too specialized to have been the ancestors of the bony fish.
The bony fish (Osteicthyii)
Bony fish are by far the most numerous of all fish, comprising about 24,000 living species, and they come in an amazing variety of forms and sizes (ranging from a half-inch-long sea horse weighing a fraction of an ounce to a 1,000 pound blue marlin). The purported evolutionary relationship of all these fish is at best highly speculative.
All bony fish have gills for breathing and fins for swimming. Starting from front to back, the most important fins for swimming are the paired pectoral fins (which are typically attached to the posterior margin of the skull), the generally smaller paired pelvic fins (that occupy a position near the anus), and the caudal fin (tail fin).
Bony fish are divided into two groups, the lobe-finned fish, known mostly from fossils, and the vastly more numerous ray-finned fish. Both have fins made up of bony rays, but the lobe-fins have fin rays mounted on a short, fleshy stalk supported by successive segments of bone.
It is the superficial resemblance of these bony fins to tetrapod legs that has led evolutionists to speculate that the lobe-fin fish are the ancestors of tetrapods in the late Devonian (approximately 380 million years ago). So, let’s focus our investigation on the lobe-fins.
The lobe-fin fish (Sarcopterygii)
The lobe-finned fish have been divided into two rather dissimilar groups, the Dipnoi (lungfish) and the Crossopterygii (coelacanths and fossil relatives).
Lungfish (Dipnoi)
There are only three surviving types of lungfish. They are all eel-like in appearance, and have long and slender fleshy pectoral and pelvic fins, which are highly mobile. This group derives its name from the fact that they have air sacks (“lungs”) that function at least partially in breathing (though all, at least in their immature state, have functional gills as well). The fact that these fish can breathe air, survive out of water for long periods of time, and have the ability to pull themselves along on their bellies (i.e. “walk”) across mud flats with the aid of their fins, has caught the imagination of some evolutionists who consider them to be ancestral to tetrapods.
Many living fish are air-breathers and “walkers”
But air-breathing fish are not uncommon among living fish species. For example, many popular aquarium fish (such as the paradise fish, betta and gourami) are surface air-breathers that can actually drown if kept under water! Evolutionists are not even in agreement on whether lungs evolved before gills (as proposed by the famous vertebrate evolutionist Alfred Romer), or gills evolved before lungs.
Even the sort of “walking” that lungfish engage in is not uncommon among living fish species. Many fish are known to pull themselves along on their bellies, with the help of their pectoral fins, across large expanses of mud flats and even dry land. For example, the northern snakehead and the “walking catfish” (Clarias batrachus) are air-breathing fish that can travel overland for considerable distances. The mudskippers are fish that breathe oxygen through their skin and “skip” along on land with the aid of their fleshy fins—indeed some of the larger species are said to skip faster than the average person can run! The climbing perch (Anabas testudineus) not only breathes air and “walks” on land but is even said to be capable of climbing trees! Yet none of these curious fish are considered by evolutionists to be ancestors of tetrapods—they are simply interesting and specialized fish. In fact there are even “flying fish” (with specialized fins that permit them to fly or glide in the air for hundreds of yards over water), but evolutionists have never considered them to be ancestors of birds.
Crossopterygians
Most evolutionists now look to fossil Crossopterygians for the ancestors of tetrapods—even though none of them are known to be capable of either walking or breathing out of water.
The distinguishing features of these fish are the division of the skull into anterior and posterior units (considered similar to embryonic tetrapod skulls); and fleshy pectoral fins containing bony elements (considered similar to tetrapod legs). These similarities have prompted evolutionists to confidently declare that Crossopterygians evolved into tetrapods.
According to evolutionists, the Crossopterygians flourished during the middle to late Devonian (extending from 385 million years ago to 365 million years ago) and all were once believed to have become extinct about 80 million years ago (even before the extinction of the dinosaurs).2
The coelacanth—one of many “living fossils”
However, in 1938 a fishing trawler netted a strange large blue fish in the Indian Ocean off the coast of Madagascar. This distinctive fish was soon identified as a Crossopterygian fish previously known only from the fossil record as the coelacanth.
Coelacanths are distinctly different from all other living fishes. They have an extra lobe on their tails (compared to other lobe-finned fish) and are the only living animal to have a fully-functional joint in their cranium, which allows the front part of the head to be lifted when the fish is feeding.
The discovery of a coelacanth came as a surprise to evolutionists. (It was comparable to finding a living dinosaur, because these fish were believed to have become extinct 80 million years ago when they disappeared from the fossil record.) However, since 1938, dozens of living coelacanths have been found and studied, some as far as 7,000 miles away from the location of the first sightings!3
Understandably, evolutionists are puzzled by how coelacanths could disappear for over “80 million years” and then turn up alive and well in the twentieth century. They speculate that the fossilized coelacanths lived in environments favoring fossilization, whereas modern coelacanths live at great depths (over 600 feet) in caves and overhangs of steep marine reefs that don’t favor fossil formation. This, however, is special pleading, since essentially no modern marine environment favors the formation of fossils and, indeed, none are being formed, as this would require rapid burial, which is not observed under normal conditions.
More importantly the coelacanth (along with many other “living fossils”) show that evolutionists can never assume that a plant or animal did not live during any particular period of assumed geologic time simply because it does not appear in the fossil record of this period. If 200-pound coelacanths can “hide” for “80 million years,” it would seem anything can hide.
Another reason finding a living coelacanth caused so much surprise at the time of its discovery was that coelacanths were widely believed to be the ancestors of the tetrapods. Indeed, many evolutionists assumed that the very reason the coelacanth disappeared from the fossil record was because they evolved into land-dwelling tetrapods; yet here they were very much alive—and swimming!
Coelacanths don’t walk
At the very least, evolutionists expected to observe some hint of walking behavior in the coelacanth, but sadly the fish have done nothing to accommodate them. Although living coelacanths have often been observed swimming in their natural habitat, they have never been observed walking. Indeed, coelacanths have been observed swimming backward, upside-down, and even standing on their head! Alas—they absolutely refuse to walk on land or in the sea.
Evolutionists look to other lobe-fins
Since living lobe-fin fish have not met expectations, evolutionists have turned to other fossilized lobe-fins for the ancestors of tetrapods. (After all, one can speculate endlessly about fossils without fear of contradiction—until they turn-up alive.)
Currently, the three most popular Crossopterygian candidates for ancestors of tetrapods are Eusthenopteron, Panderichthys, and the recently discovered Tiktaalik.
Eusthenopteron
For several years, the evolutionist’s “gold standard” of fish with “legs” has been the fossil fish Eusthenopteron (which, like the coelacanth, has fleshy pectoral fins with bones). If you have seen an artist’s illustration in a textbook showing a fish walking out of the water, most likely it was Eusthenopteron.
Like most other jawed fish, Eusthenopteron has its pectoral fin girdle (bones that anchor the pectoral fins) attached to the back of its skull by means of a dermal bone called the cleithrum. Dermal bones develop directly from connective tissue cells under the skin, rather than from cartilage models as is the case for endochondral bones. (Fish scales, by the way, are dermal bones as well, and reside just under the superficial layer of the skin.)
Panderichthys
Panderichthys is yet another fossil Crossopterygian fish that has been declared to be an ancestor of tetrapods. Panderichthys lacks dorsal and ventral fins and has a relatively small tail fin (thus looking less obviously fish-like than Eusthenopteron).
Like the other Crossopterygian fish, Panderichthys has thick bony pectoral fins. Evolutionists argue that the shape of these fins and their pectoral girdle look more like that of tetrapods than Eusthenopteron. But Daeschler, Shubin, and Jenkins—the discoverers of Tiktaalik—claim that “Panderichthys possesses relatively few tetrapod synapomorphies, and provides only partial insight into the origin of major features of the skull, limbs and axial skeleton of early tetrapods.” As a result, they insist that “our understanding of major transformations at the fish-tetrapod transition has remained limited.”4
Tiktaalik to the rescue?
In the April 2006, issue of Nature, Daeschler, et al. reported the discovery of several fossilized specimens of a Crossopterygian fish named Tiktaalik roseae. These well preserved specimens were found in sedimentary layers of siltstone—cross-bedded with sandstones—in Arctic Canada.4
Like the other lobe-fin fish, Tiktaalik was declared to be late Devonian (between 385-359 million years old) by means of a “dating” method known as palynomorph biostratigraphy. This method presumes to date sedimentary rock layers on the basis of the assumed evolutionary age of pollen and spores contained in the rock. Most importantly, the discoverers of Tiktaalik claim that it “represents an intermediate between fish with fins and tetrapods with limbs.”
Tiktaalik is a fish
Whatever else we might say about Tiktaalik, it is a fish. In a review article on Tiktaalik (appearing in the same issue of the scientific journal Nature that reported the discovery of Tiktaalik), fish evolution experts, Ahlberg and Clack concede that “in some respects Tiktaalik and Panderichthys are straightforward fishes: they have small pelvic fins, retain fin rays in their paired appendages and have well-developed gill arches, suggesting that both animals remained mostly aquatic.”5
In other respects, however, Ahlberg and Clack argue that Tiktaalik is more tetrapod-like than Panderichthys because “the bony gill cover has disappeared, and the skull has a longer snout.” The authors weakly suggest that the significance of all this is that “a longer snout suggests a shift from sucking towards snapping up prey, whereas the loss of gill cover bones probably correlates with reduced water flow through the gill chamber. The ribs also seem larger in Tiktaalik, which may mean it was better able to support its body out of water.”
Without the author’s evolutionary bias, of course, there is no reason to assume that Tiktaalik was anything other than exclusively aquatic. And how do we know that Tiktaalik lost its gill cover as opposed to never having one? The longer snout and lack of bony gill covers (found in many other exclusively-aquatic living fish) are interpreted as indicating a reduced flow of water through the gills, which, in turn, is declared to be suggestive of partial air-breathing—but this is quite a stretch. Finally, what does any of this have to do with fish evolving into land dwelling tetrapods?
Are the pectoral fins of Tiktaalik really legs?
Before we get into Tiktaalik’s “legs,” it might be instructive to consider an old trick question. If we call our arms “legs,” then how many legs would we have? The answer, of course, is two legs—just because we call our arms “legs” doesn’t make them legs. The same might be said of the bony fins of Crossopterygian fish—we may call them “legs” but that doesn’t necessarily make them legs.
Shubin et al. make much of the claim that Tiktaalik’s bony fins show a reduction in dermal bone and an increase in endochondral bone.6 This is important to them because the limb bones of tetrapods are entirely endochondral. They further claim that the cleithrum (a dermal bone to which the pectoral fin is attached in fish) is detached from the skull, resembling the position of the scapula (shoulder blade) of a tetrapod. They also claim that the endochondral bones of the fin are more similar to those of a tetrapod in terms of structure and range of motion. However, none of this, if true, proves that Tiktaalik’s fins supported its weight out of water, or that it was capable of a true walking motion. (It certainly doesn’t prove that these fish evolved into tetrapods.)
The limbs of tetrapods
The limbs of tetrapods share similar characteristic features.
The limbs of tetrapods share similar characteristic features. These unique features meet the special demands of walking on land. In the case of the forelimbs there is one bone nearest the body (proximal) called the humerus that articulates (flexibly joins) with two bones, the radius and ulna, further away from the body (distal). These in turn articulate with multiple wrist bones, which finally articulate with typically five digits. The hind limbs similarly consist of one proximal bone, the femur, which articulates with two distal bones, the tibia and fibula, which in turn articulate with ankle bones; and finally with typically five digits. In order to support the weight of the body on land, and permit walking, the most proximal bones of the limbs must be securely attached to the rest of the body. The humerus of the forelimb articulates with the pectoral girdle which includes the scapula (shoulder blade) and the clavicle (collar bone). The only bony attachment of the pectoral girdle to the body is the clavicle.
The femur of the hind limb articulates with the pelvic girdle, which consists of fused bones collectively called the pelvis (hip bone). It is this hind limb—with its robust pelvic girdle securely attached to the vertebral column—that differs radically from that of any fish. (The tetrapod arrangement is important for bearing the weight of the animal on land.)
All tetrapod limb bones and their attachment girdles are endochondral bones. In the case of all fish, including Tiktaalik, the cleithrum and fin rays are dermal bones.
It is significant that the “earliest” true tetrapods recognized by evolutionists (such as Acanthostega and Ichthyostega) have all of the distinguishing features of tetrapod limbs (and their attachment girdles) and were clearly capable of walking and breathing on land. The structural differences between the tetrapod leg and the fish fin is easily understood when we realize that the buoyant density of water is about a thousand times greater than that of air. A fish has no need to support much of its weight in water where it is essentially weightless.
The fins of fish (including Tiktaalik)
Essentially all fish (including Tiktaalik) have small pelvic fins relative to their pectoral fins. The legs of tetrapods are just the opposite: the hind limbs attached to the pelvic girdle are almost always more robust than the fore limbs attached to the pectoral girdle. (This is particularly obvious in animals such as kangaroos and theropod dinosaurs.) Not only are the pelvic fins of all fish small, but they’re not even attached to the axial skeleton (vertebral column) and thus can’t bear weight on land.
While the endochondral bones in the pectoral fins of Crossopterygians have some similarity to bones in the fore limbs of tetrapods, there are significant differences. For example, there is nothing even remotely comparable to the digits in any fish. The bony rays of fish fins are dermal bones that are not related in any way to digits in their structure, function or mode of development. Clearly, fin rays are relatively fragile and unsuitable for actual walking and weight bearing.
Even the smaller endochondral bones in the distal fin of Tiktaalik are not related to digits. Ahlberg and Clack point out that “although these small distal bones bear some resemblance to tetrapod digits in terms of their function and range of movement, they are still very much components of a fin. There remains a large morphological gap between them and digits as seen in, for example Acanthostega: if the digits evolved from these distal bones, the process must have involved considerable developmental rearranging.”
So is Tiktaalik a missing link?
Finally, what about the popular claim that Tiktaalik is the “missing link” between fish and tetrapods?
In their review article on Tiktaalik, Ahlberg and Clack tell us that “the concept of ‘missing links’ has a powerful grasp on the imagination: the rare transitional fossils that apparently capture the origins of major groups of organisms are uniquely evocative.” The authors concede that the whole concept of “missing links” has been loaded with “unfounded notions of evolutionary ‘progress’ and with a mistaken emphasis on the single intermediate fossil as the key to understanding evolutionary transition.”5
Sadly, “unfounded notions” of this kind continue to be uncritically taught and accepted in the popular media and in our schools. Even more sadly, these unfounded notions have been used to undermine the authority of Holy Scripture.

Footnotes
- Wilford, John Noble, Fossil Called Missing Link From Sea to Land Animals, The New York Times, Late Edition—Final, Section A, Page 1, Column 5, April 6, 2006.
- New Fossils Fill the Evolutionary Gap Between Fish and Land Animals, http://www.nsf.gov/news/news_summ.jsp?cntn_id=106807, 2006.
- Another was recently caught near Indonesia.
- Daeschler, Edward B. , Shubin, Neil H., and Jenkins, Farish A., A Devonian tetrapod-like fish and the evolution of the tetrapod body plan, Nature 440(7085): 757–763, 6 April 2006.
- Ahlberg, P.E. and Clack, J.A., News and Views, Nature 440(7085): 747–749), 6 April 2006.
- Shubin, Neil H., Daeshler, Edward B., and Farish, Jr., Jenkins A., The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb, Nature 440(7085): 764–771), 6 April 2006.



Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis