
2.4 Natural Selection, Yes; Evolution, No
Darwin and Biologic Change
by
Dr. Gary Parker
on
March 28, 2016
Republished with permission and featured in
Creation: Facts of Life
(1) What does “fittest” mean?
The definition of “fittest” guarantees that natural selection must be accepted as a fact. Most people assume that “fitness” refers to features of structure, function, or behavior that suit an organism for a particular role in its environment. It doesn’t. Fitness is defined by scientists solely in relation to relative reproductive success. Members of a population that leave the most offspring to the next generation are fittest by definition.
You may have thought the dark-colored peppered moth was fittest to survive in a polluted forest because it was most camouflaged. But what if the extra melanin production interfered with, say, sex hormone production and made the dark-colored moths sterile? Obviously, the superior camouflage would not make such a moth fittest to survive! Evolutionists think the camouflage helped, of course, but the dark moths were really determined to be “fittest to survive” because a greater percentage of their offspring survived in polluted forests than the percentage for any other color form.
Think about zebras. Their survival depends on their ability to outrun lions. So, the fastest zebra would be fittest, right? Not necessarily. Suppose the fastest zebra was hard of hearing or had a poor sense of smell. It could have outrun the lion and the rest of the herd—if only it had sensed the lion’s coming! Or suppose the fastest zebra had bones that broke easily, poor digestion, and/or caught diseases easily. What looks fit to us superficially may not turn out to be fittest in nature.
So, the only way to determine fitness is to make notes on organisms in the first generation, wait for the struggle for survival to take place, then see which organisms actually left the most offspring to the next generation. To see how scientists calculate fitness, let’s work through Figure 12, a simple example involving one pair of genes, A and a, which produce three varieties of organisms: AA, Aa, and aa. These gene combinations (genotypes) could be used to represent a variety of traits (phenotypes), e.g., tall-medium-short, fast-medium-slow, red-pink-white, smart-average-dull, heavy-medium-light, etc.
We’ll start the first generation with 100 individuals: 50 AA, 30 Aa, 20 aa. The second generation coming through the struggle for survival includes 20 AA, 60 Aa, and 20 aa. All other things being equal, it’s already obvious that organisms with genotype Aa were fittest, winning the struggle for survival, since they’re the only group that increased in numbers. The numerical fitness of each group can be easily calculated. First, divide the number in the second generation in each category by the number in the first; that gives 20/50 = 0.4 for AA; 60/30 = 2.0 for Aa; and 20/20 = 1.00 for aa.
Note the highest survival ratio is the 2.0 for the Aa fittest, or winners, in this example. Calculate the standardized fitness value by dividing each “survival ratio” by the highest (2.0 for Aa here). This last step always gives the winner a fitness value of 1.00 and ranks other groups from 0 (a loser with no survivors) to some fraction of 1.00. The aa fitness here is 1.0/2.0=0.5, meaning the aa’s survived about “half as well” as the fittest Aa’s. The AA’s did worst at surviving, about “20 percent as well” as the fittest (20/50 = 0.4 and 0.4/2.0 = 0.2).
Several profound and often misunderstood consequences follow from the simple calculation of fitness:
| A. Static Population | B. Increasing Population | C. Decreasing Population | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trait categories | AA | Aa | aa | Tot. | AA | Aa | aa | Total | AA | Aa | aa | Tot. |
| First generation, G1 | 50 | 30 | 20 | 100 | 50 | 30 | 20 | 100 | 50 | 30 | 20 | 100 |
| Second generation, G2 | 20 | 60 | 20 | 100 | 40 | 120 | 40 | 200 | 10 | 30 | 10 | 50 |
| G2/G1 ratio | 20/50 | 60/30 | 20/20 | 40/50 | 120/30 | 40/20 | 10/50 | 30/30 | 10/20 | |||
| “Numerical fitness” | 0.4 | 2.0 | 1.0 | 0.8 | 4.0 | 2.0 | 0.2 | 1.0 | 0.5 | |||
| H, highest ratio (survival winner) | 2.0 | 4.0 | 1.0 | |||||||||
| Ratio G2/G1 ÷ H | 0.4/2 | 2/2 | 1/2 | 0.8/4 | 4/4 | 2/4 | 0.2/1 | 1/1 | 0.5/1 | |||
| “Standardized fitness” | 0.2 | 1.0 | 0.5 | 0.2 | 1.0 | 0.5 | 0.2 | 1.0 | 0.5 | |||
| Fittest | 1.0 | 1.0 | 1.0 | |||||||||
(a) “Survival of the survivors.” The definition of fitness is grounded ultimately in reproductive success, so it is sometimes called differential reproduction or net relative reproductive efficiency. In far less than pompous-sounding phrases, what that boils down to is survival of the survivors. Now you can see why natural selection, or survival of the fittest, is a fact. How is it determined which organisms will be “naturally selected” as fittest? Wait for the struggle for survival to play out from one generation to the next, then count who survived in greatest numbers! An organism may be ugly, slow, or stupid, but if its offspring survive in greatest numbers, it’s the fittest! (That may comfort some of us, as well as the opossum!)
Notice that natural selection is NOT some awesomely powerful scientific theory that enables scientists to predict future changes in populations. “Natural selection” is really just a high-sounding, misinforming term applied to the observation that some organisms in a varied population survive in greater numbers than others do—survival of the survivors. After scientists observe which organisms are “fittest” (i.e., survived in greatest relative numbers), then they can begin to speculate on why. Was it camouflage, speed, intelligence, fecundity (having lots of offspring easily), disease resistance, some combination or none of these, or just “blind luck”? Ecclesiastes 9:11 says, “The race is not to the swift, nor the battle to the strong [in our fallen world] . . . but time and chance happeneth to them all” (ASV).
Natural selection is a fact because it’s a tautology or truism, a form of circular reasoning. It is argued that the fittest are those that survive in greatest relative numbers and those that survive in the greatest relative numbers are defined as the fittest. That’s definitely true, but it’s really just an observation, not a profound theory, and begs the question of what makes some organisms fitter than others.
The story is told of a student walking to school who saw in the grass a mouse that remained absolutely motionless as a hawk soared overhead. When she asked her teacher why, the teacher explained that mice which ran were seen and killed by the hawk, so natural selection produced those which remained motionless. The next day, the student saw a mouse running to its burrow as a hawk soared overhead. When she asked her teacher why, the teacher explained how mice that remained motionless were easy targets for the sharp-eyed hawk that killed and ate them, so natural selection favored survival of the mice which ran. The “nice” thing about “survival of the survivors” is that it can explain anything: why mice run or stay put, why some species (e.g., horseshoe crabs) never changed in “600 million years” while others changed rapidly and quickly (e.g., an insect-eater thought to have evolved into horses, whales, and bats in less than “5 million years”). The so-called “proof” that natural selection produced evolution is too often merely the argument that survivors survived!
(b) Natural selection versus ecological competition. Most people just assume “natural selection” for the “fittest” means the selected variety must be increasing. Actually, natural selection has nothing to do with whether a species as a whole is increasing or decreasing in numbers or staying the same (static or stable). Look back at the calculation of fitness in Figure 12. In case A, the population was static or stable; the second generation had 100 individuals like the first one did. Now imagine the population doubled to 200, and the second generation contained 40 AA, 120 Aa, and 40 aa. What would the new fitness values be? The winner (“fittest”) being “naturally selected” is still Aa, and its reproductive efficiency is 120/30 = 4.0, which is the highest value. That means the standardized fitness of Aa, 4.0/4.0, is 1.00, the maximum value, just as it was in the static population. The fitness values for the other two groups are also exactly the same in the expanding population as they were for the static case. The reproductive efficiency for aa is 40/20 = 2.0, so its standardized fitness is 2.0/4.0 (the “winning” efficiency) = 0.5, “one-half” the maximum, as before. The numbers for AA are 40/50 = 0.8, and 0.8/4.0 = 0.2, exactly 20 percent of maximum as in the static population.
What if the species population is decreasing? Who’s the fittest then? Imagine the population declined by half, and the second generation was 10 AA, 30 Aa, 10 aa (50 total). Again, Aa is the best survivor or fittest, this time because it declined the least in population. Aa’s numbers are 30/30 = 1.0, and 1.0/1.0 (the highest) is 1.0. The aa’s again did “half” as well: 10/20 = 0.50, and 0.5/1.0 = 0.5. The AA “losers” got a fitness score of “20 percent” maximum, just as before: 10/50 = 0.2, and 0.2/1.0 = 0.2. Notice, however, the species population is decreasing dramatically. In this case, being the “fittest” only means being the high scorer on the losing team!
Being the fittest, then, is no guarantee of survival at all. It may only mean you are likely to be the last of your kind to die out! Fitness has to do with competition within a group; survival of the group often depends on competition among different groups, often related to changing environmental factors—loss of habitat, increase or decrease in temperature or moisture levels, changes in the saltiness of aquatic and soil environments, catastrophes like fires, floods, earthquakes, underwater landslides, etc. So, for example, it’s NOT natural selection that determines whether the dull and sluggish opossum or the sleek and daring cheetah survives; it’s ecology, interaction among different groups and the environment (and so far the opossum is outscoring the cheetah in the ecological competition!).
(c) Intraversus interspecific competition. Many people have the mistaken notion that natural selection involves, for example, competition between lions and zebras. Not at all. Natural selection is NOT lion versus zebra; it’s lion versus lion (which can catch a zebra) and zebra versus zebra (which can escape the lion). In other words, natural selection is NOT INTERspecific competition (between species); it’s INTRAspecific competition (within species). By analogy to humankind, natural selection is competition among classmates and friends for dates on a Saturday night and jobs at McDonald’s, or competition among brothers and sisters for family favors. Natural selection is the ultimate sibling rivalry, a struggle to the death among members of the same species. Even members of a plant species compete with one another (not consciously, of course) for water and minerals from the soil and a place in the sun. Some variants of a species are more likely to leave more offspring to the next generation than others, but at most the intraspecific competition of natural selection produces variation within kind, NOT change from one kind to another. Natural selection, yes; evolution, no.
A classic lab kit sold to demonstrate natural selection does nothing of the sort. The kit includes two different species of flour beetle, Tribolium confusum and T. castaneum. By changing temperature and moisture conditions and adding predators and different hiding places, students can see one beetle species survives better under this condition, the other beetle species under that. Competition between different species as conditions change is ecological competition, not at all natural selection among members of the same group.
Evolutionists, however, did report an example of natural selection that once occurred in a flour beetle experiment. A mutant beetle occurred in one species, and offspring of that beetle eventually wiped out other members of that species—natural selection in action. The supposedly “new and improved” beetle species then lost the ecological competition with the other beetle species under conditions that the pre-mutant beetle species formerly won. As evolutionists recognize, winning the natural selection battle can lead to losing the ecological war—“mischievous results” of natural selection one evolutionist called it.
(d) Succession versus evolution. Evolution is a hypothetical process that is supposed to change a few simple forms over time into many complex and varied forms. There is a real process of change through time in which a few life forms are followed by a series of more and more complex and varied forms, but the real process is ecological succession, NOT evolution. If you watched an area of bare rock over time, as farmers and scientists have, you could observe a series of changes from lichens to moss, ferns, shrubs, and trees, but the lichens didn’t evolve into the moss, nor the moss into the ferns, etc. Rather, each living community changed the environment in ways that paved the way for the next community to move in. (Plants “move” by scattering spores and seeds which sprout when conditions are right.) Lichens can break down rock, producing enough soil for mosses. Mosses build more soil, and hold moisture, paving the way for shrubs. Shrubs break up the rock further, anchor the soil, and provide shade to decrease moisture loss, paving the way (in the proper climate) for trees.
As the plant communities change, so do the animals. Protozoans are followed successively by worms, insects, birds, and mammals. Existing species from another area move in as conditions become favorable—ecology, not evolution. It is migration of different kinds, NOT mutation of one kind into others, that produces ecological succession. Succession involves only tens or hundreds of years, NOT millions.
Death is not a necessary part of ecological succession, and at least some kinds in early (pioneer) communities survive through various seral stages into the final (climax) community. Lichens grow on bare rock, for example, but lichens also grow on tree bark in climax forests. Ecological succession on a global scale would have followed both creation (“multiply and fill”) and the Flood (migration from Ararat).1 As discussed later, dramatic environmental changes caused by the Flood would favor both (a) selection for different adaptations among pre- and post-Flood members of the same kind, and also (b) survival of different kinds in different proportions in the pre- and post-Flood ecologies.
Ironically, natural selection and ecological competition don’t really provide adequate explanation for presumed evolutionary changes, but they do help explain changes important in the creation model.
(e) Long term versus short term advantage. Richard Dawkins, Great Britain’s leading spokesman for evolution, refers to evolution by natural selection as the “blind watchmaker.”2 In contrast to creation by plan and purpose looking toward a goal, natural selection, Dawkins asserts, is a “blind” process that does not plan, has no purpose, and can’t look ahead toward goals. Natural selection is merely opportunistic, rewarding chance combinations of traits with a slight advantage in Darwin’s ceaseless “war of nature.”
Dawkins is right about natural selection, but wrong about the nature of the living world. Natural selection cannot plan ahead; selection is only the observation that certain trait combinations will win the immediate struggle for survival, becoming, by definition, the fittest—no matter what that does to the future of the species. That can have a devastating impact on living things, the exact opposite of the evolutionist’s hopes and dreams.
Consider territorial population control.3 Many birds and mammals regulate their population through a series of complex instincts and “ritualistic combat” in which no death occurs and no predators are necessary. Sea lions, for example, limit their population by “allowing” breeding only on certain restricted territories on small beaches. Males who fail to stake out a territory one year must wait until later years to breed. That guarantees plenty of food for the species as it cruises the Pacific. Suppose a chance mutation knocked out the instinct for territorial recognition. Such a mutant male might establish a new breeding colony on another island and pass on his unrestricted urge to breed. Descendants of such a male would automatically win the struggle for survival in the short term, but the long term effects might include over-hunting their range and even bringing the species to extinction—or at least replacing gentle territorial control with harsher predatory control. Indeed, some evolutionists blame a large percentage of extinction on the exploitation of environmental resources automatically rewarded by natural selection, which is “blind” to long term consequences.
Natural selection tends to favor specialists, and that also produces problems long term. In a given environment, specialists are usually more efficient at exploiting food sources than generalists, and evolutionists recognize the tendency for natural selection to convert generalized ancestral populations into ever more specialized descendants. When the environment changes, highly adapted, specialized varieties tend to lose out to the adaptable, generalized forms—if there are any left. Again, natural selection seems to promote short-term survival at the expense of long-term extinction. As we shall see in the chapter on fossils, the long-term survivors over and over again are the generalized, adaptable forms like those God created to multiply and fill the earth, not the specialized forms natural selection generated to exploit short-term advantage.
Dawkins is right about the blindness and failure to plan by natural selection, but that makes him wrong about evolution and the history of life on earth.
(f) Brake or accelerator? Remember, evolution may not be true, but natural selection is. Natural selection is a process at work in our fallen world; it is a description of what happens when different varieties of the same gene-trading species compete for limited resources. As we have seen, the results of natural selection in action are often the opposite of what evolutionists expected, and the exact opposite of what the public is told.
Calling natural selection “survival of the fittest” conjures up an image of a positive, progressive process. Natural selection really operates as the “great eliminator” or “terminator,” and might be better called “unsurvival of the unfittest.” Think back on the famous peppered moth case. Natural selection did NOT produce a “new and improved moth”; the dark moth was already present. Pollution made the light form less camouflaged, and so (presumably) natural selection eliminated more light than dark moths. Had natural selection “gone to completion” and totally eliminated the light moth, the species might now be well on the road to extinction, since reduction in pollution has now made the light moth more camouflaged again.
Note also that natural selection only promoted increased death of less camouflaged moths; it did nothing to produce either dark or light color. Mutations are supposed to produce new traits for selection to select, but known mutations are either neutral (having no effect) or harmful, producing defects, disease, and disease organisms. Perhaps the most important role of natural selection in a fallen world (corrupted creation) is acting as a brake, slowing down the accumulation of harmful mutations, eliminating or reducing genetic decay by producing “unsurvival of the unfittest.”
All scientists agree that elimination of the unfit is a major consequence of natural selection in our present world, but a process that works at best to make tomorrow no worse than today is no process for producing the evolutionist’s dream of upward, onward progress. Eliminating defects to repair an old car may keep it running, but it will never turn a mini-van into a Formula 1 race car!
(g) Fitness versus adaptation. Adaptations are features and functions that suit an organism for its roles in its environment. Fitness is determined by counting survivors in Darwin’s “war of nature;” adaptation is determined by engineering or design analysis. A woodpecker is admirably designed for drilling holes in wood, regardless of how well it is surviving. Professional evolutionists freely admit that fitness and adaptation are quite different concepts determined in quite different ways,4 but that major difference is almost always overlooked in popular nature programs and children’s literature, and is often ignored in introductory college biology textbooks. Professional evolutionists do believe that at least some of the time well-adapted organisms should show greater fitness: i.e., leave more offspring to the next generation than their competitors. Creationists already know, of course, that organisms were created with adaptations for survival so they could multiply and fill the earth.
There is no convincing evidence or argument that fitness or natural selection lead to adaptation, but there is ample evidence and logic for the reverse: adaptation can lead to natural selection! If organisms already have certain adapted or adaptable traits, then, as they multiply over the earth, they will more likely survive as the “fittest” and be “naturally selected” in some environments rather than others. In his article on “Adaptation” in the Scientific American book Evolution, Lewontin5 emphasizes this point over and over again:
. . . evolution cannot be described as a process of adaptation because all organisms are already adapted. . . .
. . . adaptation leads to natural selection, natural selection does not necessarily lead to greater adaptation. . . .
That is, adaptation has to come first, before natural selection can act. Natural selection obviously cannot explain the origin of traits or adaptations if the traits have to be there first.
Lewontin recognizes that this simple (but crucial) point is often overlooked, so he gives an example. As a region becomes drier, he says, plants can respond by developing a deeper root system or a thicker cuticle (waxy coating) on the leaves, but “only if their gene pool contains genetic variation for root length or cuticle thickness” (emphasis added). Here again, the genes for deep roots and thick, waxy coats must be present among the genes of a kind before natural selection can select them. If the genes are already there, we are talking only about variation within kind, i.e., creation, not evolution. As creationists were saying even before Darwin’s time, natural selection does not explain the origin of species or traits, but only their preservation—how and where certain varieties survive as they multiply and fill the earth.
Lewontin is an evolutionist and outspoken anti-creationist, but he honestly recognizes the same limitations to natural selection that creation scientists do:
. . . natural selection operates essentially to enable the organisms to maintain their state of adaptation rather than to improve it (emphasis added).
Natural selection does not lead to continual improvement (evolution); it only helps to maintain features that organisms already have (creation). Lewontin also notes that extinct species seem to have been just as fit to survive as modern ones, so he adds:
. . . natural selection over the long run does not seem to improve a species’ chances of survival, but simply enables it to “track,” or keep up with, the constantly changing environment (emphasis added).
Natural selection works only because each kind was created with adaptations (design features) and sufficient variety to multiply and fill the earth in all its ecologic and geographic variety. Without realizing it at the time, Darwin actually discovered important evidence pointing both to God’s creation (adaptation and variation) and to the corruption of creation (struggle and death).
The seven points above are all logical limits to extrapolating the hypothetical process of evolution (macroevolution) from the observable process of natural selection. It really looks like using natural selection to “reach” evolution is like using a bicycle to reach the moon; the barriers are insurmountable, no matter how much time you take. Evolutionists face two even more serious difficulties in trying to explain evolution as a result of natural selection: “compound traits” and the “origin” of new traits.
(2) Compound traits or “irreducible complexity”
Many believe any genius Darwin had is found in explaining how all the complex and varied structures and functions of living things could be produced one step at a time by the process of natural selection. Imagine you are standing at the bottom of the Empire State Building. Getting to the top looks impossible, especially if you have to do it in one huge jump. Then someone shows you the stairway. What looked like an impossibility now seems like a certainty. The climb may be long and hard, but you could make it from the bottom to the top if you took one step at a time. That’s the way most people now look at the world of living things. Producing life without the outside help of a Creator once seemed impossible. Now, say the evolutionists, the production of all life forms from simple beginnings is a virtual certainty—IF AND ONLY IF each feature is produced slowly and gradually, one step at a time.
Darwin himself, however, recognized that adaptations in living systems often depend on many parts working together simultaneously, and Darwin called such features “difficulties with the theory.” Such compound traits, or systems of irreducible complexity, are considered the most powerful argument against Darwinism and have fostered the burgeoning growth of the “Intelligent Design” (ID) movement among secular scientists today.6 Remember, natural selection can be used to turn the impossible into the highly probable IF AND ONLY IF each step in the development of an adaptation has survival value, allowing it to increase in numbers relative to its competitors.
Throwing dynamite into the fire started by Michael Denton (Evolution: A Theory in Crisis, 1985)7 and Phillip Johnson (Darwin on Trial, 1991),8 biochemist Michael Behe brought popularity to the Intelligent Design (ID) movement among secular scientists with the publication of his book (Darwin’s Black Box, 1996),9 describing stunning examples of irreducible complexity found in the “molecular machinery” of living cells: the astonishing rotary motor of the bacterial flagellum, photoreceptor/effector systems (“eyes”), complex stimulation/inhibition interactions in blood clotting and the immune system, etc.!!! Right now, let’s look at examples of compound traits on a larger scale.
Perhaps the biggest problem for evolutionists is “the marvelous fit of organisms to their environment.” As I mentioned in the first chapter, many adaptations involve whole groups of traits working together, and none of the individual pieces has any survival value (“Darwinian fitness”) until the whole set is functioning together. Remember the woodpecker? Let’s look at another example.
Since death entered the world, there are many large, predatory fish that roam the oceans. As they feed on smaller fish and shrimp, their mouths begin to accumulate food debris and parasites. Lacking recourse to a toothbrush, how is such a fish going to clean its teeth?
For several kinds of fish, the answer is a visit to the local cleaning station. These are special areas usually marked by the presence of certain shrimp and small, brightly colored fish, such as wrasses and gobis. Often fresh from chasing and eating other small fish and shrimp, a predatory fish may swim over to take its place in line (literally!) at the nearest cleaning station. When its turn comes, it opens its mouth wide, baring the vicious-looking teeth.
You might suspect, of course, that such a sight would frighten off the little cleaner fish and shrimp. No, into the jaws of death swim the little cleaners. Now even a friendly dog will sometimes snap at you if you try to pick off a tick, and it probably irritates the big fish to have a shrimp crawling around on its tongue and little fish picking parasites off the soft tissues of the mouth. (Try to imagine shrimp crawling around on your tongue!) But the big fish just hovers there, allowing the cleaners to do their work. It even holds its gill chambers open so that the shrimp can crawl around on the gill filaments, picking off parasites!
At the end of all this cleaning, the second “miracle” occurs. You might think the fish would respond, “Ah, clean teeth; SNAP, free meal!” But, no. When the cleaning is done, the big fish lets the little cleaner fish and shrimp back out. Then the big fish swims off—and begins hunting again for little fish and shrimp to eat!
The fantastic relationship just described is called cleaning symbiosis. Perhaps you have seen cleaner fish in a major public aquarium, or seen pictures of their behavior in television footage or nature magazines. Cleaning symbiosis is a well-known example of mutualism, an intimate relationship of benefit to both types of species involved, in this case, the “cleaner and the cleanee.”
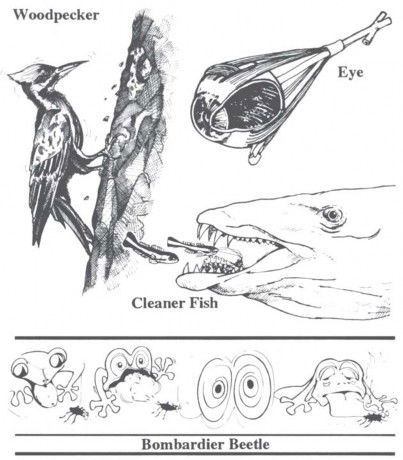
Figure 13. Darwin included structures requiring many interdependent parts in a chapter titled “Difficulties with the Theory.” Before it can have any survival value, every part of a bombardier beetle’s “cannon” must be in place, and the same is true for the woodpecker’s set of “drilling tools” and the “nerve wiring” for cleaner-fish behavior. Evolutionist Lewontin says such “perfection of structure was,” and I say is, “the chief evidence of a Supreme Designer.”
Obviously, cleaning symbiosis has survival value for both types of species involved, but does survival value explain the origin of this special relationship? Of course not. It makes sense to talk about survival value only after a trait or relationship is already in existence. Question: Did the survival value of this cleaning relationship result from time, chance, and struggle, or from plan, purpose, and special acts of creation?
The major problem is using Darwinian fitness to explain traits with many interdependent parts when none of the separate parts has any survival value. There’s certainly no survival value in a small fish swimming into a large fish’s mouth on the hope that the big fish has somehow evolved the desire to let it back out! Sea creatures don’t provide the only examples of cleaning symbiosis, either. A bird, the Egyptian plover, can walk right into the open mouth of a Nile crocodile—and walk back out again, after cleaning the croc’s mouth! On an evolutionary basis, each cleaning relationship would have to be explained separately on the basis of time, chance, struggle, and death, operating on variants of each species involved. Remember, natural selection can help explain the origin of compound traits one step at a time IF AND ONLY IF each separate step has survival value on its own.
The situation is even more dangerous for the famous “bombardier beetle.” The bombardier is an ordinary-looking beetle, but it has an ingenious chemical defense mechanism. Imagine: Here comes a mean ol’ beetle-eater, a toad, creeping up behind the seemingly unsuspecting beetle. Just as he gets ready to flash out that long, sticky tongue, the beetle swings its cannon around, and “boom!” It blasts the toad in the face with hot noxious gases at the boiling point of water, and coats the toad’s tongue with a foul-tasting residue. Now that doesn’t actually kill the toad, but it surely kills its taste for beetles! Pictures show the toad dragging its tongue across the sand trying to get rid of the foul taste.
Successful firing of the bombardier beetle’s cannon requires two chemicals (hydrogen peroxide and hydroquinones), enzymes, pressure tanks, and a whole series of nerve and muscle attachments for aim and control. Try to imagine all those parts accumulating by time, chance, and natural selection. One crucial mistake, of course, and “boom!” the would-be bombardier beetle blows itself up, and there’s surely no evolutionary future in that! Trial and error can lead to improvement only if you survive the error!
Creationists and evolutionists agree that adaptations such as the woodpecker’s skull, cleaning symbiosis, and the bombardier beetle’s cannon all have survival value. The question is, how did they get that way: by time, chance, struggle, and death, or by plan, purpose, and special acts of creation? When it comes to adaptations that require several traits all depending on one another, the more logical inference from the evidence seems to be creation.
(3) Origin of Traits
Darwin’s theory also points us back to creative acts when it comes to the origin of traits. In spite of the title of his book, Origin of Species, the one thing Darwin never really dealt with was the origin of species. That is, he never explained the origin of the truly new traits needed to produce a truly new kind of organism, something more than just a variation of some existing kind. There are many other logical limits to extrapolation from natural selection to evolution, but the simplest is this: natural selection cannot explain the origin of traits.
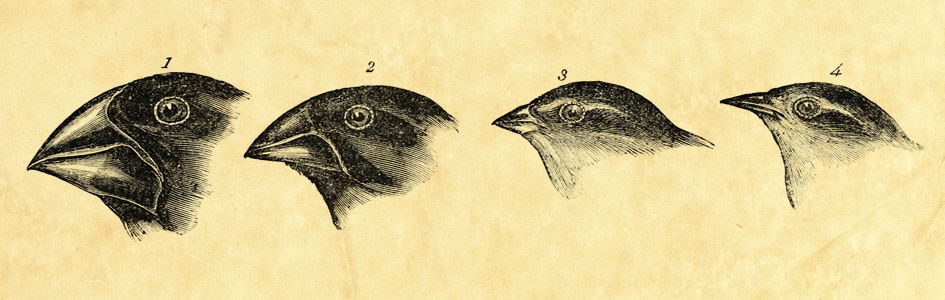
Take the famous example of “Darwin’s finches” (Figure 14). On the Galapagos Islands, Darwin observed a variety of finches, some with small beaks for catching insects, others with large beaks for crushing seeds, and one with the ability to use spines to pry insects from their tunnels. How did Darwin explain the “origin” of these various finches? Exactly the same way a creationist would. He saw finches with variation in beak type on the South American mainland and presumed these finches might have reached the islands on a vegetation mat or something similar. The ones with seed-crushing beaks survived where seeds were the major food source, and those with insect-catching beaks out-reproduced others where insects were the major source of food. Given finches with a variety of beak types, then, natural selection helps us to explain how and where different varieties survived as they multiplied and filled the earth. That, of course, is just what a creationist would say—except that a biblical creationist would add that the “struggle and death” part of migration did not begin until man’s rebellion ruined the world God had created without death. (Contrast Genesis 1–2 with the Fall in Genesis 3.)

Figure 14. “Darwin’s Finches.” Darwin explained the location of (a.) Pangenesis: Use and Disuse. Darwin called natural selection “the preservation of favored races,” and he recognized that selection alone could not explain origin. When it came to the actual origin of new traits, Darwin wrote that it was “from use and disuse, from the direct and indirect actions of the environment” that new traits arose. About 40 years before Darwin, a famous French evolutionist, Jean Lamarck, argued for this kind of evolution based on the inheritance of traits acquired by use and disuse. Most books on the subject hint that we should laugh at Lamarck—but Darwin believed exactly the same thing.
Consider the supposed origin of the giraffe. According to both Darwin and Lamarck, the story begins back on the African prairies a long time ago. Because of prolonged drought, the prairie dried up. But there were green leaves up in the trees, and some of the animals started stretching their necks to reach them. As a result, their necks got a little longer (Figure 15). Now that could be partly true. If you really work at it hard enough and long enough, you could add a little bit to your height. People used to do that to get into the army or some special service where you have to be a certain height. The problem, however, is that the offspring of “stretched” parents start off just as small as all the others. The long neck could not be passed on to the next generation.

Figure 15. For the origin of new traits, Darwin (like Lamarck) resorted to “use and disuse” and the inheritance of acquired characteristics. Giraffes got longer necks, for example, because their ancestors stretched for leaves in trees, then passed on more neck “pangenes” to their offspring. This idea of “progress through effort” contributed to the early popularity of evolution, but has since been disproved.
Like others of his time, Darwin didn’t know about the mechanism of heredity. He thought that each organ produced ‘‘pangenes’’ that would collect in the blood and flow to the reproductive organs; so, a bigger neck made more neck pangenes. Some people still believe this sort of concept. You’ve probably run into people who say, for instance, that people will eventually have bigger heads because we think a lot, and no toes because we wear shoes all the time. Darwin even used pangenes to “explain” why (in his opinion) wives grew to resemble their husbands as both got older.
It seems people knew as little about giraffes in Darwin’s time as they did about heredity. Because their neck is so long, there’s a huge distance between a giraffe’s heart and its brain. It needs auxiliary pumps to get blood to the brain so it won’t faint when it raises its head up—and it needs pressure reducers so that when it bends its head down to take a drink, it won’t blow its brains out! A long neck without these features would be deadly.
Science has since disproved these “flimsy facts” of early evolutionary thought, but back in Darwin’s time, pangenes captured people’s imagination probably even more than natural selection did. To some, Darwin’s original theory of evolution suggested continual progress. How do you make something happen? By use and disuse. If you want to get smarter, use your brain, and both you and your children will be smarter. If you want to be strong, use your muscles, and not only will you get stronger, but so will your children.
Well, almost unfortunately, that’s not the modern theory of evolution. The use-disuse theory didn’t work and had to be discarded. The modern evolutionist is called a neo-Darwinian. He still accepts Darwin’s ideas about natural selection, but something new (neo-) has been added. The modern evolutionist believes that new traits come about by chance, by random changes in genes called “mutations,” and not by use and disuse.
(b.) Mutations. Almost everyone has heard about mutations—from Saturday morning cartoons or horror movies, if nowhere else. In those flicks, some atomic disaster produces people with gnarled skin, one big bulging eye, and other “new traits.” In the real world, mutations are responsible for a number of genetic defects, including hemophilia (bleeders’ disease), loss of protective color in the skin and eyes (albinism), and certain kinds of cancer and brain malfunction.
We have abundant evidence that various kinds of radiations, errors in DNA replication, and certain chemicals can indeed produce mutations, and mutations in reproductive cells can be passed on to future generations. Figure 16 shows some of the changes that have been brought about in fruit-fly wings because of mutations: shorter wings, very short wings, curled wings, spread-apart wings, miniature wings, wings without cross veins. Students in my genetics classes work with these fruit flies each year, crossing different ones and working out inheritance patterns.
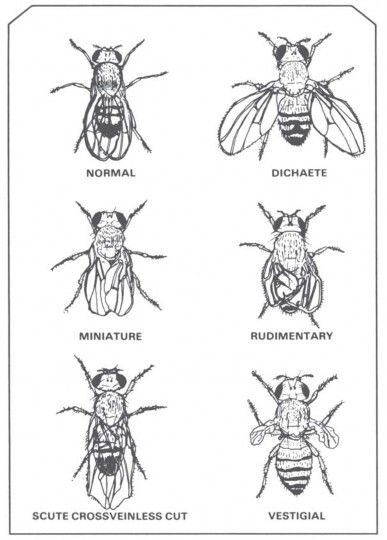
Figure 16. Mutations are random changes in genes (DNA), often caused by radiation. The mutations in the wings above were produced by x-raying fruit flies. According to the modern, neo-Darwinian view, mutations are the source of new traits for evolution, and selection culls out the fittest combinations (or eliminates the “unfittest”) that are first produced just by chance. Mutations certainly occur, but are there limits to extrapolating from mutational changes to evolutionary changes (e.g., “fish to philosopher”)?
Then there’s the flu virus. Why haven’t we yet been able to solve the flu problem? Part of the problem is that this year’s vaccine and your own antibodies are only good against last year’s flu. (They don’t usually tell you that when you get the shot, but it’s already out of date.) The smallpox virus has the common decency to stay the same year in and year out, so once you’re vaccinated or build up an immunity, that’s it. The flu virus mutates quite easily, so each year its proteins are slightly different from last year’s. They are still flu viruses, but they don’t quite fit our antibodies, so we have to build up our immunity all over again. When it recombines with animal viruses (on the average of once every ten years), the problem is even worse.
Mutations are certainly real. They have profound effects on our lives. And, according to the neo-Darwinian evolutionists, mutations are the raw material for evolution.
Is that possible? Can mutations produce real evolutionary changes? Don’t make any mistakes here. Mutations are real; they’re something we observe; they do make changes in traits. The question remains: do they produce evolutionary changes? Do they really produce new traits? Do they really help to explain that postulated change from molecules to man, or fish to philosopher?
Creation: Facts of Life

Dr. Parker, a leading creation scientist and former AiG speaker, presents the classic arguments for evolution used in public schools, universities, and the media, and refutes them in an entertaining and easy-to-read style. A must for students and teachers alike! This is a great book to give to a non-Christian as a witnessing tool.
Read OnlineFootnotes
- Gary Parker, Exploring the World Around You (Green Forest, AR: Master Books, 2002).
- Richard Dawkins, The Blind Watchmaker (New York: W.W. Norton, 1986).
- Parker, Exploring the World Around You.
- Dobzhansky, Ayala, Stebbins, and Valentine, Evolution.
- Richard C. Lewontin, “Adaptation,” Scientific American (and Scientific American book Evolution) (September 1978).
- Jonathan Wells, Icons of Evolution (Washington, DC: Regnery Pub., 2000).
- Michael Denton, Evolution: A Theory in Crisis (London: Burnett Books, 1985).
- Phillip Johnson, Darwin on Trial (Washington, DC: Regnery Gateway, 1991).
- Michael Behe, Darwin’s Black Box (New York: The Free Press, 1996).




Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis