
1.4 Comparative Similarities: Homology
Evidence of Creation?
by
Dr. Gary Parker
on
February 13, 2016
Republished with permission and featured in
Creation: Facts of Life
If God made people as people, why are we full of “animal parts”? Look at your arm for a moment and try to picture the bones inside. There’s one bone attached to the body, two bones in the forearm, a little group of wrist bones, and bones that extend out into the fingers. As it turns out, there are many other living things that have forelimbs with a similar pattern: the foreleg of a horse or dog, the wing of a bat, and the flipper of a penguin, for example, as shown in Figure 6. Biologists use the term “homology” for such similarities in basic structure.
Why should there be that kind of similarity? Why should a person’s arm have the same kind of bone pattern as the leg of a dog and the wing of a bat? There are two basic ideas. One of these is the evolutionary idea of descent from a common ancestor. That idea seems to make sense, since that’s the way we explain such similarities as brothers and sisters looking more alike than cousins do. They have parents closer in common.
Using descent from a common ancestor to explain similarities is probably the most logical and appealing idea that evolutionists have. Some think that our ability to classify plants and animals on a groups-within-groups hierarchical basis virtually forces scientists to treat evolution as a “fact.” However, we can classify kitchen utensils on a groups-within-groups basis, but that hardly forces anyone to believe that knives evolved into spoons, spoons into forks, or saucers into cups and plates.
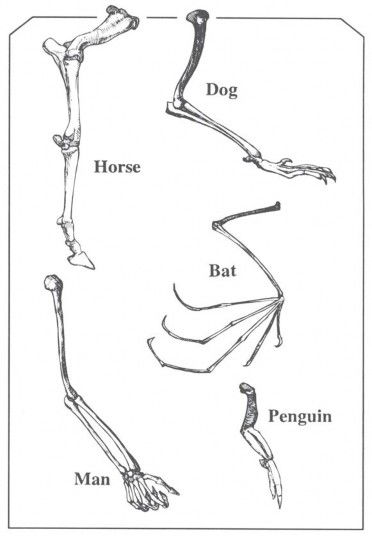
Figure 6. Bones in the human arm, the forelimbs of horses and dogs, a bat’s wing, and a penguin’s flipper all share a similarity in basic structural pattern called homology. What does this similarity (homology) mean: descent from a common ancestor (evolution), or creation according to a common plan (creation)?
After all, there’s another reason in our common experience why things look alike. It’s creation according to a common plan. That’s why Fords and Chevrolets have more in common than Fords and sailboats. They share more design features in common.
What’s the more logical inference from our observation of bone patterns and other examples of homology: descent from a common ancestor, or creation according to a common plan? In many cases, either explanation will work, and we can’t really tell which is more reasonable. But there seems to be times when the only thing that works is creation according to a common design.
I get support for my claim again from Denton,1 in his chapter titled “The Failure of Homology.” Dr. Denton is not only a research scientist with a Ph.D. in molecular biology, but also an M.D. with an intimate knowledge of comparative anatomy and embryology. He admits his desire to find naturalistic explanations for patterns of similarity among organisms (homology), but he also admits the failure of evolutionary explanations.
Like every other scientist, Denton recognizes the striking similarity in bone pattern evident between vertebrate fore- and hindlimbs. Yet no evolutionist, he says, claims that the hindlimb evolved from the forelimb, or that hindlimbs and forelimbs evolved from a common source. I was once taught to refer to corresponding parts of the male and female reproductive systems as “sexual homology.” Homology, in that case, could not possibly be explained by descent from a common ancestor; that would mean that males evolved from females, or vice versa, or that human beings evolved from some animal that had only one sex.
Worse yet for evolution, structures that appear homologous often develop under the control of genes that are not homologous. In such cases, the thesis that similar structures developed from genes modified during evolutionary descent is precisely falsified.
In frogs, for example, the five digits on each limb grow out from buds on the embryonic paddle; in human embryos, the digits form as the tissue between them is resorbed. Here quite different gene-enzyme mechanisms produce similar (homologous) patterns. Structures in adult lobsters and crayfish are so similar (homologous) that the same lab instructions can be used for dissecting either, yet the crayfish egg develops directly into the adult form while the lobster egg reaches the homologous pattern through a free-swimming larval stage.
Our observation of similarity or homology is real enough, but that’s true, Denton points out, “whether the causal mechanism was Darwinian, Lamarckian, vitalistic, or even creationist” (emphasis added). Although the evidence is not as spectacular and compelling as the biomolecular data, I would say the weight of our present knowledge of homology favors Denton’s final alternative: creation according to a common design.
Perhaps the clearest anatomical evidence of creation is “convergence.” The classic example is the similarity between the eyes of humans and vertebrates and the eyes of squids and octopuses. Evolutionists recognize the similarity between the eyes easily enough, but they’ve never been able to find or even imagine a common ancestor with traits that would explain these similarities. So, instead of calling these eyes homologous organs, they call them examples of “convergent evolution.” Rather than evolution, however, we have another example of similarity in structure that cannot be explained as evolutionary descent from a common ancestor.
Convergence, in the sense of similar structures designed to meet similar needs, would be expected, of course, on the basis of creation according to a common design. As we’ll see later, both the octopus eye and the vertebrate eye are complete, complex, and totally distinct from one another right from their first appearance in the fossil sequence. Biologist Michael Land2 sounds like a creationist when he mentions in passing that the vertebrate eye “shares design features but not evolution” with the eye of the cephalopod mollusks such as the octopus.
The real focus of Land’s article, however, is “divergence,” the occurrence of quite distinct structures in plants and animals that otherwise are supposed to be close evolutionary relatives. Certain shrimp-like animals that live in deep ocean darkness, he says, have compound eyes with lenses all arranged to focus light at a common point (rather than forming multiple images, as most compound eyes do). But, he continues, some members of the group have “lens cylinders” that smoothly bend the incoming light (because of smoothly varied refractive indices), whereas others have square facets with a “mirror system” for focus (utilizing even a double-corner bounce). Ingenious use of physics and geometry should be evidence enough of creation it seems to me—but there’s more.
Comparing the mirrors with the lens cylinder system, Land says, “Both are successful and very sophisticated image-forming devices, but I cannot imagine an intermediate form [or common ancestral type] that would work at all.” The kind of design in these eyes, he says, seems impossible to explain as a result of evolutionary relationship. So Land goes on to suggest that the shrimp-like animals with different systems should not be classified as evolutionary relatives, even though they are otherwise quite similar.
Even more interesting is Land’s statement about how he felt when he was trying to figure out the mirror system. He said he was “trying not to come to the conclusion that these eyes had been put there by God to confuse scientists.” They may confuse evolutionists, but may I suggest instead that these eyes were put there by God to inform scientists. As such cases show, a mind open to examples of created order can hasten and enrich the scientific search for understanding.
Some evolutionists admit they have failed to find good evidence of evolution in comparing large structures, so they are looking instead for homology among molecules. In a foundational book basically describing the three-dimensional structures first known for proteins, Dickerson and Geis3 state that “from the perfection of protein sequence and structure analysis . . . . We can pin down with great precision the relationships between the species and how the proteins evolved.” Then, with every example they give, they proceed to disprove that evolutionary prediction.
Consider hemoglobin, for example, the protein that carries oxygen in red blood cells. Dickerson says that hemoglobins pose “a puzzling problem. Hemoglobins occur sporadically among the invertebrate phyla [the animals without backbones] in no obvious pattern.” That is, they don’t occur in an evolutionary branching pattern. I would suggest that they do occur in a creationist mosaic or modular pattern, like bits of blue-colored stone in an artist’s mosaic. We find hemoglobin in nearly all vertebrates, but we also find it in some annelids (the earthworm group), some echinoderms (the starfish group), some mollusks (the clam group), some arthropods (the insect group), and even in some bacteria! In all these cases, we find the same kind of molecule—complete and fully functional. As Dickerson observes, “It is hard to see a common line of descent snaking in so unsystematic a way through so many different phyla. . . .”
If evolution were true, we ought to be able to trace how hemoglobin evolved. But we can’t. Could it be repeated evolution, the spontaneous appearance of hemoglobin in all these different groups independently, asks Dickerson? He answers that repeated evolution seemed plausible only as long as hemoglobin was considered just red stuff that held oxygen. It does not seem possible, he says, that the entire eight-helix folded pattern appeared repeatedly by time and chance. As far as creationists are concerned, hemoglobin occurs, complete and fully functional, wherever it is appropriate in the Creator’s plan, somewhat like a blue-colored tile in an artist’s mosaic.
Mosaic refers here to a picture or mural formed of many little bits of colored stone. According to the mosaic concept of kind, God used several different genes or gene sets over and over again in different combinations and proportions to make a variety of life forms, somewhat like an artist might use several different kinds of colored stones over and over in different proportions and arrangements to make a variety of artistic designs. The different bits of stone in the artist’s mosaic would correspond to the many different genes or gene sets in God’s “mosaics,” which are the various forms of living things.
According to this mosaic concept, also called modular or matrix, God used a basic plan in making living creatures, somewhat similar to the plan He used in making different non-living substances. All the countless chemical substances in the universe are made from different combinations and proportions of only about a hundred different elements, usually displayed in a “chemistry mosaic” called the periodic table. Each kind of chemical compound can be represented by a formula expressing the number, kind, and arrangement of elements within it.
Perhaps God used genes as “elements” in making the various kinds of life, so that conceivably each different kind of life could be represented by a “formula” representing the number, kind, and arrangement of different genes in its chromosomes. Such formulas would, of course, be much larger and much more complex than those for the most complicated chemical substances. Nevertheless, the mosaic concept does suggest that all the incredible variety and diversity of life forms we see about us may be constructed using only the information in a few thousand DNA segments, compared to about 100 chemical elements. Even more exciting, creationists might be able to use a mosaic pattern (or mathematical matrix) to predict the existence of unknown organisms and their features, like Mendeleev used his periodic table to predict the existence and properties of elements before their discovery.
The mosaic, non-branching (non-evolutionary) pattern of trait distribution produces practical problems for the biologist. Algae are usually classified into major groups on the basis of their pigment (greens, reds, browns, goldens, etc.), for example. But then both their structural complexity (unicellular, colonial, multicellular) and type of sexuality (iso-, hetero-, or oo-gamy) must be re-evolved independently (“convergently”) on different branches of the evolutionary tree based on color. If they are classified by level of structural complexity, then neither the color pattern nor type of sexuality can be traced back to one common ancestor. Similarly, the evolutionary tree based on type of sexuality contradicts the branching trees predicted by pigment and structural complexity.
While he was yet the internationally respected senior paleontologist at the British Museum, Colin Patterson4 stunned the scientific world by calling evolution an “anti-theory” that generates “anti-knowledge”—a concept full of explanatory vocabulary that actually explains nothing and that even generates a false impression of what the facts are.
Patterson said that he finally awoke, after having been duped into taking evolutionism as revealed truth all his life, to find that evolutionary theory makes bad systematics (the science of classification). He then proceeded to examine the data as a creationist would, in simple recognition that creationists produce testable hypotheses, and that he could understand and explain what inferences creationists would draw from the data, without either agreeing or disagreeing with them. What a superb example of healthy scientific skepticism! Patterson was able to see the data regarding homology in their wholeness, and experience the unbridled freedom to wonder not only how but whether evolution occurred!
Some are hoping that DNA comparisons and gene sequencing (“molecular homology”) can somehow salvage evolutionary classification.5 Is there anyone who hasn’t heard that DNA comparison suggests something like 98 percent similarity between man and chimpanzee? The evidence so convinced one evolutionist debater that he told the audience if a chimp asked to take his daughter out on a date, he was not sure he could say “No.” (I hope the daughter would be allowed to say “No.”) There are even some groups pushing for the extension of U.N. human rights protection to chimps and orangutans!
It only takes a trip to the zoo, of course, to convince us that man and ape share many features, and there are unseen similarities in bone, muscle, nerve and sense organs, circulatory and digestive systems, hair, milk, etc. It should be equally obvious, however, that creatures designed by the same Creator to move, eat, breathe, etc. in similar ways would have many molecular similarities in common.
An article on “The 2% Difference” (Discover, April 2006) praises evolution and puts down intelligent design, but the author (Sapolsky) actually admits and describes key evidences noted by creation scientists over the past two decades.6 “Regulation is everything,” he says. A sidewalk, fence, patio, and house may be made of bricks that are 100 percent identical, for example, but they are arranged in different ways to serve dramatically different purposes. Sapolsky points out that the brains of man and chimp operate using “the same basic building blocks” while they achieve “vastly different outcomes,” so that in his opinion “there’s not the tiniest bit of scientific evidence that chimps have aesthetics, spirituality, or a capacity for irony or poignancy.” These awesome gaps or “qualitative distinctions” between the brains of chimps and people Sapolsky credits to a “relatively few” genes that regulate the number of brain cells (neurons) produced. Sapolsky seems to forget, of course, that a dysfunctional or diseased brain has just as many neurons as the ones we call normal, and stuffing more chips into a computer does not automatically improve it. It’s not just the number of parts that produce the great gulf between human and chimp; it’s how the parts are connected. As creation scientists have long noted, and the Bible implies, living things (and their functioning parts) are not a product of substance, but of organization. At the atomic level (“dust of the ground”), all organisms are essentially 100 percent identical; if the 2 percent difference in DNA presumed for man and chimp told the other 98 percent how to organize, the differences would be at least as vast and unbridgeable as we observe.
And there’s more. The April 2006 Discover article finally admitted what creation scientists have stressed for over 20 years: “a tiny 2% difference translates into tens of millions of AGCT differences.” Indeed, a 2 percent difference among three billion base pairs would mean about 60 million code letter differences between man and chimp. So, as creationists pointed out long ago and Sapolsky admits, “There are likely to be nucleotide differences in every single gene.” In fact, reported in 2004 studies comparing chimp chromosome 22 with its presumed counterpart on human chromosome 21 showed a DNA difference of about 1.5 percent resulted in differences of more than 80 percent among the proteins produced by those genes.7 That did not surprise creation scientists, but shocked evolutionists.
Actually, studies of molecular homology have produced major controversies within the evolutionists’ camp, since DNA trees frequently disagree with evolutionary trees based on fossils and/or on comparative anatomy. The evolutionist split is greatest when it comes to conflicting attempts (based on dubious, compounded assumptions) to use molecular homology as some sort of “evolutionary clock.” After documenting the misfit of molecular data with both of two competing evolutionary views, Michael Denton8 writes this summary (p. 306):
The difficulties associated with attempting to explain how a family of homologous proteins could have evolved at constant rates has created chaos in evolutionary thought. The evolutionary community has divided into two camps—those still adhering to the selectionist position, and those rejecting it in favor of the neutralist. The devastating aspect of this controversy is that neither side can adequately account for the constancy of the rate of molecular evolution; yet each side fatally weakens the other. The selectionists wound the neutralists’ position by pointing to the disparity in the rates of mutation per unit time, while the neutralists destroy the selectionists’ position by showing how ludicrous it is to believe that selection would have caused equal rates of divergence in ‘‘junk’’ proteins or along phylogenetic lines so dissimilar as those of man and carp. Both sides win valid points, but in the process the credibility of the molecular clock hypothesis is severely strained and with it the whole paradigm of evolution itself is endangered (emphasis added).
Denton doesn’t stop with these devastating anti-evolutionary comments (and a comparison of belief in molecular clocks with belief in medieval astrology!). He also describes data from molecular homology as a “biochemical echo of typology,” where typology is the pre-evolutionary view of classification developed by scientists on the basis of creationist thinking.
Although partial data fit too easily into conflicting branching patterns, comparative similarities and homologies don’t fit well at all onto evolutionary trees. They fit instead into hierarchical (groups within groups) categories, perhaps suggesting a multidimensional matrix (a “cube of cubes” in more than three dimensions). When Mendeleev discovered the pattern God used in creating the chemical elements, he was able to predict the existence and properties of elements not then known to science. Creationists may one day discover predictive patterns of trait distribution among living things, and prediction is the real measure of merit among scientific theories.
Creation: Facts of Life

Dr. Parker, a leading creation scientist and former AiG speaker, presents the classic arguments for evolution used in public schools, universities, and the media, and refutes them in an entertaining and easy-to-read style. A must for students and teachers alike! This is a great book to give to a non-Christian as a witnessing tool.
Read OnlineFootnotes
- Denton, Evolution: A Theory in Crisis, chapter 7.
- Michael Land, “Nature as an Optical Engineer,” New Scientist (October 4, 1979).
- Richard E. Dickerson and Irving Geis, The Structure and Action of Proteins (New York: Harper and Row, 1969).
- Colin Patterson, address at American Museum of Natural History, New York, November 5, 1981 (summarized by Gary Parker and Luther Sunderland in Acts and Facts, Impact No. 108, Institute for Creation Research, El Cajon, CA, December 1982).
- Clete W. Knaub, A Critique of Molecular Homology (Santee, CA: Institute for Creation Research, 1983), master’s thesis.
- Robert Sapolsky, “The 2% Difference,” Discover (April 2006).
- The International Chimpanzee Chromosome 22 Consortium, “DNA Sequence and Comparative Analysis of Chimpanzee Chromosome 22,” Nature (May 27, 2004).
- Denton, Evolution: A Theory in Crisis, p. 306.

Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis