

Adam and Eve
Chapter 10
Genetics Confirms the Recent, Supernatural Creation of Adam and Eve
by
Dr. Nathaniel T. Jeanson
and
Jeffrey P. Tomkins
on
April 1, 2018
; last featured
February 21, 2021
Republished with permission and featured in
Searching for Adam
Abstract
The advent of modern genetics has seen the evolutionary community redouble its efforts to argue for human-primate common ancestry and against the traditional Christian understanding of the origin of the human race. As has been argued in previous chapters, a careful reading of Genesis 1–11 indicates that God created Adam and Eve supernaturally and without prior ancestry, and that all of humanity traces their ancestry back to this original couple — and not to a group of primates or proto-humans. Combined with a careful reading of the rest of Scripture, this narrative places the creation date of Adam and Eve approximately 6,000 years ago and places another population bottleneck about 4,500 years ago at the time of the Flood. This scriptural framework leads to very specific expectations about the genetic differences among humans and other species, expectations that can be scientifically tested against modern genetic data. In this chapter, we contend that genetics confirms the recent, supernatural creation of Adam and Eve and refutes the evolutionary narrative on human origins.

This chapter is from the book Searching for Adam, available in our online store.
Overview
Since most of the data that we’re going to discuss is already present within the technical scientific literature, the purpose of this chapter is to take this relatively unknown and obscure knowledge and present it in what we hope is an understandable and accessible manner for non-geneticists. To expound the details of the genetics of human origins in great depth would require a book-length treatment. Conversely, since most of the contents of this book chapter have already been argued, defended, and published as separate technical papers, we will provide here a summary of these papers with references for the more technically minded reader to explore later.
Because the genetics of human origins is a scientifically complex issue that becomes technical very quickly, we have simplified this chapter by organizing it around four major questions:
- From whom did humans originate: ape-like primates or fully human people?
- How many individuals spawned the human race: a population or a pair?
- When did humans originate: hundreds of thousands of years ago or about 6,000 years ago (i.e., ancient or recent)?
- Where did modern human populations originate: Africa or Ararat?
Though specific elements that will be covered under each of these questions are probably more familiar to the average reader (e.g., claims like “humans are 99% genetically identical to the apes,” “human chromosome 2 is the result of a fusion,” etc.), we have chosen to take a more comprehensive view rather than an apologetic medley approach. Our intention is to demonstrate that the biblical creation model accounts, not just for a handful of select genetic observation but for the entire body of genetic evidence available today.
Introduction: A Critical Scientific Point
To recognize the strength of our conclusions in genetics, the reader needs to understand only one major technical scientific point. Surprisingly, this point is not any singular genetic observation. It is rather a careful understanding of how science works.
What follows should be uncontroversial. Since creationists and evolutionists were both taught their understanding of science from a common source — the scholarly educational community of the Western world — both agree on the specifics of how science should operate. For example, evolutionists didn’t learn their trade from creationist institutions, and we didn’t learn science in the back closet of a cloistered creationist enclave, either.1
Like many scientists, we learned our most memorable lessons on the nature and operation of science via trial and error. For example, while in a graduate course on developmental biology, my fellow students and I (Jeanson) were required to prepare short, mock grant proposals in lieu of tests. Specifically, this assignment involved writing up the proposal and then presenting it orally before a small group of students and professors.
After completing my ten-minute presentation in which I described a battery of experiments to test the scientific question in which I was interested, the professor leaned back in his chair and gave his frank assessment of my ideas. He said (paraphrased),
There are three types of experiments in the world. The first type distinguishes between two competing hypotheses, regardless of which way the experiment turns out. For example, if you hypothesize A, but the experiment demonstrates B, you’ve still learned something. This is the best and rarest type of experiment. The second type is valuable only if the experiment turns out one of the two possible ways. For example, if you hypothesize A, but the experiment does not support A and instead supports a whole host of alternative hypotheses, you’ve learned very little. If, instead, the experiment had confirmed hypothesis A, it would have been valuable.
He then said that I had proposed the third type of experiment — one in which nothing is learned regardless of the experimental outcome. Essentially, a type-3 experiment tests none of the hypotheses in question, including the one that the investigator has proposed. I had made a major — but memorable — error.
What my professor didn’t say is also critically important. Implicit in the professor’s description of my proposal was an assumption that experiments were actually going to be performed. If, instead of proposing a battery of experiments, I had simply asserted that my hypotheses were true, I would have been failed rather quickly. Stating hypotheses as fact is the cardinal sin of science, so much so that it doesn’t even receive a type designation. In fact, it’s not even in the domain of science. It’s pseudoscience.
For example, consider the question of what molecule is the substance of heredity, the instruction manual for building our physical features during the process of development. If we claim that “vital forces and biorhythms from Jupiter” are the real substance, and if we perform zero experiments to test or reject our claim, we’re simply spouting pseudoscience (and we would probably be laughed at by most intelligent human beings).
Instead, if we hypothesize that a chemical molecule called DNA is the substance, we have a hypothesis we can test. Another investigator might hypothesize that protein, not DNA, is the substance of heredity. If we try to test these hypotheses by analyzing the biochemical composition of sperm and egg, we would discover that we performed a type-3 experiment — sperm and egg possess both DNA and protein, which reveals nothing about which substance carries the hereditary information.
However, if we had discovered that sperm and egg lacked one of the two substances, we would have performed a type-2 experiment — the result would have eliminated one of the hypotheses, but it would not have positively confirmed the other (after all, there might be many hypotheses on what substances control heredity, and these hypotheses would need to be eliminated as well). To perform a type-1 experiment, we would have had to show that only DNA — and not protein — was the substance of heredity.
These sorts of experiments were done in the early part of the last century. In these experiments, investigators used organisms that were easy to work with, such as bacteria and viruses. Since some viruses infect bacteria by injecting certain chemical substances into their hosts that allow the virus to propagate itself, investigators found themselves with an elegant experimental system. In other words, if scientists could figure out what exactly the virus injected, they would know what the substance of heredity was in these organisms.
Since proteins contain certain chemicals (e.g., sulfur) that DNA lacks, and since DNA contains certain substances that proteins lack (e.g., phosphorus), chemically labeling sulfur in one experiment and phosphorus in the other would distinguish between these two hypotheses. When the viruses grown in the presence of chemically labeled sulfur were allowed to infect bacteria, the sulfur (e.g., protein) stayed on the outside of the bacteria. By contrast, when the viruses grown in the presence of chemically labeled phosphorus were allowed to infect bacteria, phosphorus (e.g., DNA) was found inside the bacterial cells. Furthermore, when the investigators analyzed the offspring of the viruses, these offspring contained chemically modified phosphorus — but not chemically modified sulfur. Clearly, the substance of heredity was DNA — and not protein.
We have to ask if the claim is simply a bold assertion of fact or if it is actually based on a scientific test.
Hence, to evaluate origins claims, we first have to determine if a claim is in the realm of science. In other words, we have to ask if the claim is simply a bold assertion of fact or if it is actually based on a scientific test. If it is based on the latter, we can proceed with determining which category of experiment the claim represents. Claims that represent type-3 experiments have no further relevance to the origins debate. In contrast, type-2 and type-1 tests have the potential to uncover something new about the competing origins hypotheses, but only type-1 experiments rigorously test young-earth creation (YEC) and evolution head-to-head (Table 1).
| Experiment Type | Models Compared | Ramifications | Frequency in Origins Debate |
|---|---|---|---|
| 1 | Creation vs. Evolution | The only head-to-head test in the origins debate | Rare |
| 2 | Evolution vs. itself (or Creation vs. itself) | Useful in refuting one of the models; useless in confirming a model | Occasional |
| 3 | No models compared | Completely useless in the origins debate | Very frequent |
Evolutionists agree with the essence of what we’ve just described.2 This agreement is borne out both historically and presently. Historically, one of the most common criticisms of the creation model is that it falls in the realm of pseudoscience — that it doesn’t make experimentally testable predictions but, instead, makes bald assertions of fact. Presently, in its promotion of theistic evolution (or as they say, evolutionary creation) the BioLogos community continues to repeat this accusation:
The reason Christian anti-evolutionary approaches are absent from the mainstream scientific literature is not because scientists are theologically or philosophically biased against them, but rather because they offer little in the way of useful tools for making accurate predictions about the natural world.3 [emphasis added]
Thus, all origins positions can agree that testable, accurate predictions are critical to science, and the ability of creationists and evolutionists to make them will be the major focus of this chapter.
We also intend to demonstrate that creationist predictions are scientifically superior to those of evolutionists.
However, while evolutionists agree with the nature of science as we described above, we intend to illustrate how evolutionists of all stripes fail to practice it — on each of the four major arenas of scientific investigation on the question of human origins (from whom, how many, when, and where humans originated) — and that, in contrast to the assertion above, creationists do make accurate predictions about the natural world and about human origins in particular. We also intend to demonstrate that creationist predictions are scientifically superior to those of evolutionists.
I. From Whom: Ape-like Primates or Fully Human People?
When considering human origins, the most natural place to start is on the question of whether humans have an ape-like ancestry. Before we can discuss the minutiae of the genetics of the human race, we need to ask whether our race is indeed human or whether we are simply highly evolved primates. Ever since Darwin, evolutionists have claimed that apes represent our closest living biological relatives.4 Evolutionary creationists (a.k.a. theistic evolutionists) agree and expect to find unequivocal genetic evidence of a common genealogical heritage between mankind and the orangutans, gorillas, and chimpanzees. Current evolutionary literature identifies the chimpanzee as the closest living relative of humans, and evolutionists place the split between these two lineages (from a common ape-like ancestor, not a chimpanzee) about 3 million to 13 million years ago.5
In contrast, a plain reading of Scripture reveals a starkly different narrative on human ancestry. As has been argued in an earlier chapter, Genesis 1–2 teaches that God created man in His own image, categorically distinct from any animals, and that He did so supernaturally by forming Adam from the dust and Eve from Adam’s side. Human evolution from pre-existing ape-like creatures is not compatible with the Genesis narrative.
Furthermore, the rest of Scripture identifies Adam and Eve as the sole progenitors of the entire human race, and Noah, his wife, his three sons, and their wives as the most immediate ancestors of modern humans.6 Shortly after the global Flood of Noah’s day, the human ancestors of the modern “races”7 or ethnic groups formed as a result of the confusion of languages at Babel (Gen 11:8–9).8 Apes as precursors to humans do not enter the picture under the creation view.
Because of the nature of the genetic discussion that follows, the time element of creation is also critical to the ancestry question. Under the young-earth creation (YEC) view, Adam and Eve were created approximately 6,000 years ago, and the global Flood of Noah and the population bottleneck that followed occurred about 4,500 years ago. The Tower of Babel incident followed shortly (i.e., a couple centuries) after the Flood.9
These two strikingly different accounts — evolution and YEC — for the origin of humans lead to very different expectations about the genetics of modern humans and apes. In some cases, however, the expectations are obviously the same. For instance, from an anatomical perspective, great apes are the most similar creatures to humans, and both sides can make a general prediction that, from a genetic perspective, apes should be the most similar to humans. While humans share different levels and traits of morphological similarity with gorillas, orangutans, and chimpanzees that don’t seem to indicate any clear evolutionary pattern, the current evolutionary consensus is that humans should be most similar to chimpanzees genetically — although this widely accepted paradigm has recently been disputed based on analyses of morphological traits by several evolutionists who claim that orangutans are the closest human relative.10
As another example, both models accept the science of empirical genetic discovery. Hence, to claim that the existence of the basic science of genetics somehow validates one model over the other would be erroneous — a type-3 experiment that fails to distinguish among the competing ideas in question. Therefore, it is essential to clearly identify the specific predictions of each model in order to distinguish which genetic data actually constitute a type-1 experiment (e.g., one that differentiates YEC from evolution) and which constitute lesser types of experiments.
Are Humans 99% Genetically Identical to Chimpanzees?
One common example of a type-2 experiment is predicting the genetic difference between humans and chimpanzees. The evolutionary model has very specific expectations about this figure, and a discrepancy between predictions and facts should result in the rejection of the evolutionary hypothesis. However, since the YEC model does not make specific predictions about human-ape genetic differences, a match between evolutionary expectations and scientific fact would not inform the origins debate (i.e., would not be decisive in evolution’s favor).
But the silence of the YEC model on human-chimp genetic differences is not a weakness of the model. We could just as well challenge the evolutionists to predict the number of animals that were taken on board Noah’s ark. This request would be fruitless and irrelevant to the debate since a global Flood and an ark are not part of the evolutionary model. However, if the YEC model failed to predict the numbers on board the ark accurately, then we would need to reevaluate aspects of the YEC model. Conversely, since human-ape ancestry is not part of the YEC model, the actual number of genetic differences between humans and chimpanzees is, at best, a type-2 experiment for testing the claim that humans descended from ape-like creatures — successful evolutionary predictions would not vindicate evolution in the origins debate, while evolutionary predictive failures could be grounds to reject the evolutionary view.
With these experimental parameters in mind, we can now investigate the actual human-chimp genetic comparison in depth. If we think of genetic inheritance as analogous to copying the text of a book, the process of passing on genetic information from one generation to the next is similar to the process of transcribing the text of a book. To make the analogy tighter, inheritance is like copying the text of a book without having a perfect spell checker,11 and then using the corrupted copy as the template for the next round of copying.
Biologically, the text of the genetic book is contained in a chemical substance called DNA. The DNA in our cells is, in essence, a chemical instruction manual for building and maintaining our anatomy and physiology from conception to death. The actual instructions are encoded in a 4-letter chemical alphabet, and the combination of these letters into chemical “words” and “sentences” carries biological meaning. In total, the DNA in our cells is billions of letters long — a very large biological “book.”
When DNA is copied in sperm and egg cells prior to conception, the copying process is imperfect. The rate of copying mistakes (called mutations) has been measured in both humans and chimpanzees, and the rates are fairly similar. About 60 mutations happen each generation.12
Using rounded numbers, if the human and chimpanzee lineages split 3–13 million years ago, and if the years from one generation to the next are about 20 years, then 150,000–650,000 generations have passed since the two species last shared a common ancestor.13 In each lineage, about 60 DNA mutations happen in each of those hundreds of thousands of generations leading to an expectation that the DNA of humans and the DNA of chimpanzees should differ by about 18–80 million DNA letters.14
Thinking of DNA again like a book, we can measure book sizes by their word count, and if we wanted to be very technical, we could measure it by the total letter count. Since the total letter count in humans and chimpanzees is around 3 billion DNA letters,15 evolutionists expect about a 1–3% genetic (DNA) difference between these two species today.16
The actual difference is about 12% — a number that is about ten times higher than the predicted value.17 Though the scientist responsible for identifying this fact is a young-earth creationist, this discovery is not the result of creationist manipulation of data to fit a pre-determined conclusion. If you read the fine print in the original evolutionary publication that announced the determination of the chimpanzee DNA sequence, you can reach a similar conclusion.18 Humans and chimpanzees are not 99% identical. They are only 88% identical, which means that the two species differ by nearly 400 million (400,000,000) DNA letters!19
Thus, the question of human-chimpanzee DNA differences offers no assistance to the evolutionary model on at least three counts. First, whatever the difference is, it cannot falsify the YEC model, making it a type-2 experiment at best. Second, current evolutionary predictions for the human-chimp genetic difference fail to account for the gigantic genetic gap between these two species.
Third, the evolutionary prediction of a 1% difference isn’t really a prediction at all. The evolutionary time at which the human and chimpanzee lineages split has been revised to fit the genetic data. Earlier predictions for the time of divergence for these species were originally in the 3 to 6 million year range,20 and the measurement of the DNA copying error rate in chimpanzees caused some investigators to (controversially) bump the time back further to ~13 million years.21 Thus, the absolute difference between humans and chimpanzees isn’t a confirmed prediction as much as it is a post hoc retrofitting of predictions to facts.
These evolutionary problems aside, we are still left with the question of how to evaluate the YEC model on the human ancestry question.
These evolutionary problems aside, we are still left with the question of how to evaluate the YEC model on the human ancestry question. If human-ape genetic differences do not test validity of the YEC model of human origins, what experiment can? What genetic expectations follow from the specific YEC narrative?
In short, the answer is that, if YEC is correct, then YE creationists should be able to explain human-human DNA differences and ape-ape DNA differences [as opposed to human-ape DNA differences] without any need to reference or invoke common ancestry. In other words, YE creationists make predictions for genetic differences among individuals that share a common ancestor under the YEC view (i.e., all humans), not for individuals that were created separately (i.e., humans and apes), and these predictions can be compared to the genetic facts.
If genetic data matched these YEC expectations, would this result require rejection of the evolutionary model? Since evolutionists have spent years refining their own ideas about human-human and ape-ape genetic differences (and also believe that special creation as an alternative is unacceptable), this result would probably do nothing to settle the debate about human origins. In essence, it would be another example of a type-2 experiment — if the results are inconsistent with the YEC expectations, then perhaps the scientific elements of the YEC model should be reevaluated. But if the results confirm the YEC expectations, this discovery would probably do little to change the evolutionary claims about human-ape common ancestry.
Since subsequent sections will explore this question further, the major remaining question in this section is whether the claimed evolutionary evidences for human-ape ancestry are valid type-1 experiments. The evidences listed on the BioLogos website are presented as such — as being unequivocal proof of common ancestry and as very inconsistent with the YEC view. The evidences in the mainstream scientific literature assume the same. But is the claim true?
Relative Genetic Patterns/Nested Hierarchies
Nearly every single one of the evidences presented by BioLogos and mainstream geneticists represents a type-3 experiment or, at best, type-2. For example, one of the most common evidences cited in favor of an ape ancestry in the human lineage is the relative pattern of genetic differences between humans and apes, and between humans and other species. In short, evolutionists expect natural selection to produce a branching, tree-like pattern of genealogical relationships among the living species on this planet.22 They further expect that, if humans arose via the process of natural selection from an ape-like ancestor, then genetic comparisons among humans, apes, and other species should reveal a branching, tree-like pattern as well.
This expectation contrasts to the expectation about the percent DNA differences between humans and chimpanzees that we discussed earlier. The earlier expectation was a quantitative prediction; the current expectation is a qualitative prediction. That is, qualitatively, if humans have ancestry prior to the first Homo sapiens, then evolutionists expect humans to be relatively close genetically to the great apes, then slightly less close genetically to the rest of the primates, then even less similar genetically to other mammals, and quite different genetically from invertebrates and plants. To be clear, the absolute number of differences is not so critical as long as the same relative pattern (in this case, a nested hierarchical pattern) holds true.
For this argument to carry any scientific weight as a type-1 experiment in support of evolution, the YEC model would need to predict a different pattern. Otherwise, this argument would represent another type-3 experiment — useless to the overall origins debate.
However, it doesn’t take much reflection to see that YEC and evolution make the same prediction about the relative genetic hierarchies found in nature. Under the YEC model, God designed the entire universe, including the various kinds of biological life that exist in it, and we would expect to find that life fits a design pattern. Since humans are made in God’s image, we can get a sense for what kinds of design patterns God might have used by examining the patterns that result from human designs. Examples of nested hierarchies abound among the designed things in our world.
For example, designed means of transportation easily fit a relative hierarchical pattern. This fact is unequivocal. Sedans resemble SUVs more than they resemble tractor trailers, and all three vehicles have more in common than do sedans and amphibious assault vehicles. The latter two vehicles have more in common with one another than with submarines, and this simple pattern matches the type of hierarchy that we see in biology.23
Therefore, nested hierarchical patterns are as much the expectation of the YEC view as they are of the evolutionary view. The relative hierarchy of genetic differences among humans, great apes, mammals, and invertebrates fits the YEC model at least as well as the evolutionary one. So, to claim nested hierarchical patterns in the biological world as exclusive evidence of evolution would be analogous to claiming that the existence of people proves YEC. Neither claim constitutes a legitimate scientific experiment. Both are type-3 experiments and, therefore, reveal nothing about the validity of either view, despite the confident claims of evolutionists to the contrary.24
While these two examples (absolute and relative genetic differences between humans and the apes) do not constitute an exhaustive review of all the claimed genetic evidences for human-ape ancestry, they represent some of the most prominent, and they illustrate the Achilles’ heels of the remaining ones — failure to satisfy the requirements of a type-1 experiment.
Human Chromosome 2 Fusion?
Consider another example. If we return to our book analogy, just as the text of a book is broken up into chapters, so also the billions of letters in the DNA code for humans and chimpanzees are broken up into major divisions called chromosomes. However, because DNA comes from each parent, these chromosomes come in pairs.
Evolutionists have claimed for years that the human chromosome pair number 2 is actually an accidental fusion of two pairs of ancestral chromosomes inherited from ape-like creatures.
Evolutionists have claimed for years that the human chromosome pair number 2 is actually an accidental fusion of two pairs of ancestral chromosomes inherited from ape-like creatures.25 In short, they claim that the human-chimp ancestor had 48 chromosomes. Today, humans have 46. Since chromosomes come in two copies — e.g., the ape-like ancestor would have had 2 pairs of 24 chromosomes, and humans today have 23 pairs of chromosomes — and since humans have fewer total chromosomes than apes, evolutionists claim that one of the ancestral pairs of chromosomes fused to another ancestral pair of chromosomes. This would reduce the total chromosomes count from 48 to 46.26
Since the YEC view makes no overt predictions about the differences between humans and chimpanzees in DNA organization or in the structure of DNA, the existence of a chromosome fusion would not have said anything relevant to the human origins debate. However, in this case evolutionists also made their claim prematurely, before all the evidence was acquired. Effectively, the evolutionary claims about the structure of human chromosome 2 represented a prediction rather than an observation.
Recent reanalysis of human chromosome 2 has contradicted this evolutionary prediction. No evidence for a fusion exists. In fact, the alleged site where the fusion supposedly took place actually represents a highly organized, functional gene (in our analogy, think of genes as words or sentences).27 Thus, starting from the assumption of human-ape common ancestry, evolutionists have actually made a failed prediction about the structure and function of DNA within our cells.
The failed evolutionary prediction on chromosome function extends beyond the purported fusion site. The BioLogos community has claimed that overall arrangement of DNA along chromosomes among humans and the great apes is inexplicable apart from common ancestry: “There is no good biological reason to find the same genes in the same order in unrelated organisms, and every good reason to expect very different gene orders.”28
Do evolutionists actually have a large body of experimental results demonstrating “no good biological reason to find the same genes in the same order in unrelated organisms”? In the few cases where functional analyses have been performed, the results contradict this evolutionary assertion. The chromosomal context in which genes find themselves appears to play a significant role in how the genes function.29 In fact, human-designed computer code must also follow specific formats and contextual guidelines as well. So our previous analogy of human-designed systems as we applied to the idea of hierarchy holds true here as well. Thus, whether applied to predicted DNA differences or DNA function, the evolutionary model of common ancestry has not been vindicated.
Conversely, the prediction of function is actually one of the few arenas in the question of human ancestry in which a type-1 experiment could be conducted. Evolutionists and creationists make very different predictions about the function of the billions of DNA letters in the human sequence, and experiments testing function would clearly distinguish which model makes better predictions, as we demonstrate below.
Shared Genetic “Mistakes”?
To make the point from a different angle, the members of BioLogos have made a host of claims on their website about shared “pseudogenes” and other types of purported shared biological “mistakes” in apes and humans. In fact, two of the three main “facts” that the website lists as genetic evidence for human evolution involve an implicit statement about function.30 In reality, hardly any actual experiments have been performed on the billions of DNA letters in humans and chimpanzees. “Pseudogene” actually represents a premature label for a particular segment of DNA that resembles a broken gene but which had never been experimentally tested for function. Thus, virtually all claims that BioLogos and other evolutionists have made about genetic “mistakes” are not arguments for evolution but bald assertions without a basis in experimental fact. Technically, this would make these arguments pseudoscience. However, for the sake of discussion, we’re willing to entertain these claims as predictions stemming from the assumption that evolution is true.
Conversely, from the assumptions about human ancestry inherent to the YEC model, creationists have published a testable, predictive model of genetic function31 (see references for details). For the particular DNA differences that we examined, we expect them to function in each organism’s respective biology, whereas the evolutionary model claims that these particular DNA sequences are functionally neutral and are a reflection, therefore, of ancestry alone. Since precious few experiments have actually been done on genetic function, we now have a basis for doing a type-1 experiment in the future. By experimentally changing these sequences, we can evaluate whether or not these differences are functional — and confirm or reject the predictions of each origins model.
For other DNA sequences, a few experiments have been performed, and the trajectory is not looking good for evolution. For example, after the human DNA sequence was elucidated in 2001, it was widely proclaimed that the vast majority of our billions of DNA letters were useless, non-functional leftovers of our evolutionary heritage and therefore called “junk” DNA.32 However, scientists didn’t actually do any experimental tests on the billions of letters until the Encyclopedia of DNA Elements (ENCODE) project was initiated in 2003. The first tier of ENCODE only examined about 1% of the human genome as an initial test, and they found preliminary evidence for pervasive function for the vast majority of those billions of letters.33 Then after extending this type of research to the entire human genome, using mostly human cell lines (not fresh tissues from living humans) they reported in 2012 that at least 80% of the genome had significant levels of biochemical function.34 It wasn’t useless junk after all.
Many new discoveries in recent years are now pushing this level of functionality even higher. The leader of the ENCODE project, Ewan Birney, is predicting that the human genome will soon prove to be 100% functional.35 Needless to say, the traditional neo-Darwinian evolutionists outside the practical biomedical genetics community of ENCODE are outraged that the data is not supporting their dogmatic evolutionary claims.36
In addition to these genome-wide results, other studies focusing on specific examples of “poster child” evolutionary pseudogenes regularly damage the credibility of the evolutionary claims. For example, the beta-globin pseudogene has obvious evidence for function,37 and one of the favorite pseudogene examples (e.g., vitellogenin) of the BioLogos geneticist, Dennis Venema, can also no longer be labeled a non-functional relic.
Specifically, Venema claimed, “Humans have the remains of a gene devoted to egg yolk production in our DNA in exactly the place that evolution would predict.”38 But recent research has exposed this as nearly impossible to reconcile with the facts.39 The supposed evidence for this “egg yolk” gene is so pitiful that it’s hard to imagine how anyone could have seriously entertained this hypothesis in the first place. It’s like identifying the letter “e” in the Bible, finding the same letter in Darwin’s On the Origin of Species, and then claiming that the books were modified from a common ancestor — you really have to stretch your imagination to accept this claim. Conversely, there is so little DNA remnant of the egg yolk gene that it requires a real strain of the imagination to see why some evolutionists pursued this line of reasoning in the first place. Current data suggest that they mistook a functional DNA sequence (enhancer element) inside a genomic address messenger gene involved with brain tissue function, for a non-functional egg yolk gene “remnant.”40 Not surprisingly, the BioLogos community has downplayed the significance of these accumulating discoveries and tried to turn the tables on creationists with clever rhetorical games. Rather than admit the obvious damaging implications for evolution,41 the BioLogos staff has turned the argument around and challenged creationists to explain the remaining data that BioLogos claimed demonstrated non-function.42 In fact, Dennis Venema recently went so far as to claim, “Having the complete genome sequences for a variety of great apes makes looking for additional shared mutations a trivial exercise, and it is no exaggeration to say that there are thousands of examples that could be used.”43
The BioLogos rejoinder misses the big picture and the point.
But the BioLogos rejoinder misses the big picture and the point. First, preliminary biochemical evidence for function does not exist merely for the two examples of pseudogenes that we discussed. It exists for at least ~80% of all the pseudogenes in humans.44 And the other 20% may still yet be found to be functional in some human tissue or under some physiological condition yet to be studied . . . and there are many. That’s the catch: many noncoding RNA genes (like pseudogenes) are only expressed under certain conditions.
Second, challenging creationists to explain the remaining examples of “non-function” assumes that actual experiments have been performed that demonstrate non-function. They have not. The reality is that we have only just begun to uncover the functionality of the human genome. Consider just how many experiments would need to be performed to conclude with any sort of confidence that a particular set of DNA sequences has zero function. The number of possible scenarios in which a DNA sequence might plausibly function is now proving to be enormous. For example, in the short nine-month window of time that represents human embryonic development, a single cell turns into a fully formed baby that contains hundreds of cell types that must execute an unimaginable number of cellular tasks. Surely the developing baby calls upon enormous swaths of DNA code to execute this developmental program — and then silences or repurposes them for the remainder of its life via another type of code (a code which is being studied by investigators in a scientific field termed “epigenetics”).45 The dynamic use of DNA sequence during development is very different than the vast majority of DNA sequence use in the adult. Experimentally testing a DNA sequence during each of these unique windows of time in which sections of DNA are used and then silenced would be an enormous (and morally questionable) experiment. However, expressed RNA sequences have been analyzed in organ donors, aborted fetal tissue, and embryonic stem cells, with the latter two involving the murder of innocent babies. Nevertheless, these morbid data have only served to increase the known functionality and complexity of the human genome. In addition, until experiments are performed in living humans, which is also unethical, it is both inappropriate and scientifically uninformed to claim “non-function” for human DNA. In short, the recent decade of experimental results on human DNA sequences that demonstrate biochemical evidence for function are just the beginning of our understanding as to the complexity and function of the genome. Perhaps the most important point that can be taken from all this is the trajectory of these results — we watched the scientific community go from claiming high levels of non-function in the early 2000s to claiming evidence for nearly pervasive function just a decade later. This suggests that more experiments will only increase the percentage of human DNA sequence that performs a biological function just as the current leader of the ENCODE project is predicting. This upward trajectory does not bode well for evolution, a fact that the BioLogos community is very reticent to admit.
Neanderthal Ancestry?
On a side note, related to the question of human-ape ancestry is the question of the relationships between Neanderthals and modern humans. Interestingly, most people would be surprised to know that evolutionists consider Neanderthals to be fully human, hence they are given the technical name “archaic humans” as opposed to modern contemporary humans. An increasing number of publications claim to have recovered DNA from ancient human or human-like samples, and the comparison of these DNA samples with those of modern humans could inform the ancestry question.
Though YEC advocates and evolutionists both agree that modern humans and Neanderthals had a common ancestor (YE creationists would say that Neanderthals are post-Flood descendants of Adam and Eve), these two positions disagree on when the Neanderthals lived — tens to hundreds of thousands of years ago (evolutionary model) versus about 4,500 years or less (YEC model). Evidence for a prehistoric46 human population could add credence to the evolutionary claim that human ancestry stretches far back in time — so far back that it touches on the boundaries of an alleged divergence from an ape lineage. Time is the magical key to the evolutionary equation, despite the fact that no viable human-ape transitional forms exist in the fossil record, as discussed in a separate chapter.
Without going into great technical detail, the short answer to the question of what Neanderthal DNA implies regarding the origins issue is that Neanderthal and ancient DNA samples appear to be too degraded and often untrustworthy for use in rigorous genetic analyses.
Without going into great technical detail, the short answer to the question of what Neanderthal DNA implies regarding the origins issue is that Neanderthal and ancient DNA samples appear to be too degraded and often untrustworthy for use in rigorous genetic analyses. In addition, analyses are perpetually plagued with DNA contamination from microorganisms and modern human DNA from lab workers.47 Finally, no one knows the rate at which Neanderthal DNA changes from generation to generation — and it might change at a rate much faster than that reported for modern human individuals.48
As things stand now, the most credible research comparing Neanderthals to modern humans merely shows that their DNA is human. The dating of the bones from the sites in which Neanderthals are found are not based on DNA, but other types of spurious data, and the evolutionists are constantly changing the dates of the material found in these locations — a fact in and of itself that shows how subjective the whole process really is.
Summary
To summarize, on the question of human-ape common ancestry, all of the claimed evolutionary evidences are type-2 or type-3 experiments that fail to eliminate the main competing hypothesis, YEC (Table 2). Instead of being a minor side issue in the bigger human ancestry debate, this very poor scientific track record for evolution represents a systematic failure across the board. In nearly every type of genetic comparison that can be performed between humans and chimpanzees, the evolutionary model has made erroneous predictions (Table 3).
| Evolutionary Claim | Actual Data | Type of Experiment |
|---|---|---|
| Human-chimpanzee genetic identity is 98-99% | Actual genetic identity is only 88% (i.e., 400,000,000 DNA differences exist between the two species) | 2 |
| Humans are genetically closer to apes than to other animal species, unequivocally demonstrating common ancestry | Relative hierarchies are characteristics of design | 3 |
| Human chromosome #2 arose via fusion of two ape-like chromosomes | The purported “fusion” site is actually a functional DNA element in a human gene | 2 |
| Gene order along chromosomes has no function, therefore shared gene order demonstrates common ancestry | Gene order along chromosomes does indeed perform a function | 2 |
| Humans and chimpanzees shared genetic mistakes (e.g., pseudogenes) | Pseudogenes appear to be functional DNA elements, not mistakes | 2 |
| Humans possess the broken remnants of an ancient chicken gene (vitellogenin) | No such remnant exists; instead the “fragment” appears to be a functional DNA element | 2 |
| Type of Genetic Comparison/Analysis | Evolutionary Success or Failure? |
|---|---|
| Total DNA differences between humans and chimpanzees | Failure to predict total genetic differences (a big genetic gap separates the two species) |
| Relative genetic differences between humans and chimpanzees | Irrelevant to debate (evolutionary comparison fails to refute the YEC model, thereby making it scientifically invalid) |
| Chromosome differences between humans and chimpanzees | Failure to predict chromosome differences (no evidence for claimed fusion event) |
| Total genetic function in humans | Current scientific trajectory points toward much more function than predicted by evolution |
| Specific examples of genetic function in humans | Failure to predict functional DNA sequences (pseudogenes and chromosomal gene order were mislabeled as “non-functional”) |
In an attempt to move the discussion forward and into the realm of type-1 experiments, creationists have published a testable, predictive model of DNA function from a YEC perspective on one of the few remaining areas of DNA function that has not yet been thoroughly investigated49 (see reference for technical details). If the evolutionists are as confident in their ideas as they claim, then we invite them to publish similar predictions of genetic function, and then to do a head-to-head experiment to test both of the ideas in the laboratory. If evolutionists are unwilling to engage in the experiment that we have proposed, at a minimum, they need to propose a different type-1 experiment.
In short, on the question of human ancestry, evolutionists have a history of making erroneous scientific predictions; they have yet to articulate a genuine genetic test by which to eliminate YEC from the discussion; and their model does not look promising in light of the trajectory of experimental results in areas where evolution and YEC could theoretically be compared head-to-head.
II. How Many: A Population or a Pair?
For many years, the discussion of the number of individuals that spawned the modern human race was not accessible to science. Fossils don’t record population sizes, and the antiquity and geography of our ancestors offer little in the way of direct data on the number of individuals alive on the planet at the dawn of Homo sapiens. Only with the advent of modern genetics have scientists been able to more directly explore this question.
However, the raw genetic data say nothing about ancestral population sizes. The evolutionary conclusion that humanity arose from a large population50 rather than a pair of individuals is a consequence of the arbitrary constraints that evolutionists bring to bear on the question. Implicit in the evolutionary claims is the assumption that DNA differences can arise only via the process of copying errors (mutations) that we discussed in the previous section. In other words, under the evolutionary model, the immediate reason why you are genetically different from your parents is that you inherited DNA from each parent. However, according to evolutionary reasoning, the ultimate reason why genetic differences exist at all in the human population is mutations in the distant past.
If you insist on this evolutionary assumption and forbid the consideration of any other hypotheses on the origin of genetic differences, then you are almost forced to conclude that humanity could not have arisen from two people in the last few thousand years. Millions of DNA letter differences exist among humans (about 3–5 million per person on average, which is about 0.1% of the total human DNA sequence),51 and the measured 60 mutations per generation can’t produce this much diversity among humans in just 6,000 years, assuming that mutation rates have always been constant.
However, it doesn’t take much reflection to see that this assumption is shortsighted. Let’s apply it to the YEC model and see how well it works. If we assume, for sake of argument, that mankind did indeed arise from two supernaturally created people (regardless of how long ago it was), and if we further stipulate that genetic differences can arise only via mutations, then we would be forced to conclude that Adam and Eve did not have any genetic differences between them (aside from the X and Y chromosomes, since these are involved in specifying gender).
But this hypothetical scenario leads to some bizarre conclusions. If Adam and Eve decided to fulfill God’s command to be fruitful and multiply, they would have passed on two identical DNA sequences to their offspring. Aside from the few mutations that may have arisen (representing 0.00000001% of the billions of DNA52 letters in our cells — a negligible fraction), Adam and Eve would have basically produced copies of themselves — not slightly modified versions of themselves as we are used to observing in our own children, but identical copies of themselves. Offspring that are completely identical to parents receive a particular label in genetics: clones. Cloning as a means to fulfill the dominion mandate is a strange position to maintain. With all the debate that currently exists over the ethics of human cloning, it is somewhat disturbing to think that God instructed the first man and woman to fill the earth by this process.
A very simple alternative hypothesis resolves the conundrum and also makes straightforward scientific sense: God could have created Adam and Eve with genetic differences from the start (Figure 1). In fact, all of us possess not just 3 billion letters of DNA in our cells. With few exceptions such as red blood cells, the cells of our body possess two versions of our 3 billion letters, which means that each of our cells has 6 billion letters. Each parent passes on only 3 billion in sperm or egg, keeping the total of 6 billion letters constant across generations. Going back in time, Adam would likely have had the same cellular arrangement — two versions of his 3 billion letters — and the same would have been true of Eve.
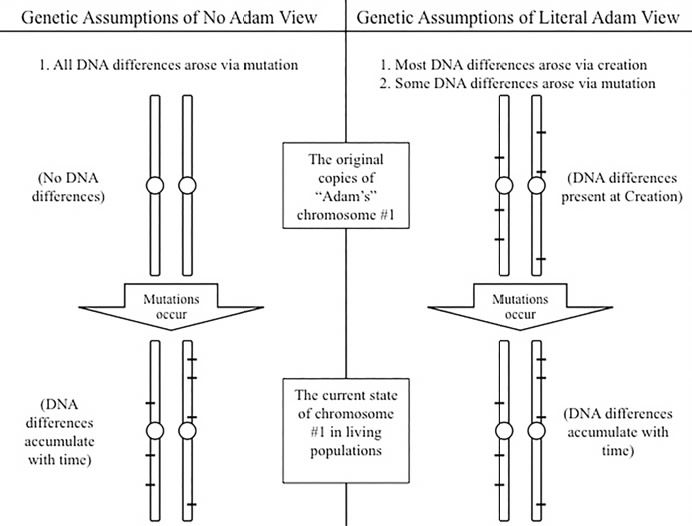
Figure 1. Fundamental assumptions about the nature of genetic change lead to very different conclusions on the original genetic state in Adam, and, therefore, on whether or not he existed in the recent past. The scenario on the left requires long periods of time to explain modern genetic diversity; the scenario on the right requires just a few thousand years to explain modern genetic diversity.
This arrangement makes sense of the DNA differences that exist in the world today. Before the Fall and after the Fall, the two different copies of Adam and Eve’s DNA would have been reshuffled via at least two processes termed recombination and gene conversion, making each offspring unique and leading to diversity within the human race. After the Fall, mutations (perhaps at a rate of 60 mutations per generation) would have occurred and added to the genetic diversity in their children,53 and leading to the production of diverse offspring (in contrast to cloning). Calculations within the parameters of this model match the worldwide DNA diversity that we observe today.54 Thus, to claim that the millions of DNA differences that separate each person from another somehow invalidates the clear teaching of Scripture about the origin of mankind from two people about 6,000 years ago is scientifically unsupportable. In fact, this type of creation model is considerably more supportive of the genetic paradigm of human diversity than the evolutionary model, as we will show.
The BioLogos website lists at least two other lines of evidence55 in support of their population-not-pair contention, but each of these falls prey to poor logic or unsound science, just like the argument above. One of the claims deals with a subsection of DNA that is repetitive in nature.56 But in attempting to explain the origin and arrangement of these sequences, the BioLogos writers assume human-ape common ancestry. Thus, as an argument against the biblical position that humans were created as a pair and distinct from the apes, it is nothing more than circular reasoning.
The second claim57 deals with the rate at which sections of DNA are swapped during sperm and egg cell production (the technical terms of two swapping processes are genetic recombination and gene conversion), but the conclusions that the BioLogos writers reach is based on erroneous assumptions and outdated science. With respect to the latter, in making their claim, the evolutionists assume only a single process of reshuffling DNA sequences (e.g., recombination) when, in fact, there are at least two (the second and, apparently, much faster process of reshuffling is gene conversion).58 Had they included this faster process in their calculations, they would have discovered that mankind’s genetic history is much shorter than they claimed.59
In summary, just like the evolutionary arguments for human-ape common ancestry, the evolutionary arguments for mankind’s origin from a large population (rather than an original pair) are nothing more than type-3 experiments, which are useless in adjudicating between creation and evolution. There is no scientific evidence that we arose from a group of individuals rather than from Adam and Eve. If evolutionists wish to continue making their claims and be taken seriously, they need to propose a type-1 experiment.
Conversely, by starting with the assumption that God created Adam and Eve with genetic diversity from the start, the YEC model can easily explain the existing genetic diversity among living humans.
Conversely, by starting with the assumption that God created Adam and Eve with genetic diversity from the start, the YEC model can easily explain the existing genetic diversity among living humans. In fact, the explanatory power of these human DNA findings is so strong that they have led to testable predictions for other species.60
III. When: Ancient or Recent?
As we’ve observed in the preceding section, using DNA sequences to function as a clock is not straightforward. In theory, just like the ticks of a clock mark off the passage of time, the transmission of another 60 DNA mutations from parent to offspring should be able to mark the passage of another generation. However, knowing how much time has passed requires knowing when the clock — whether mechanical or biological — actually started ticking. As we observed above, some (probably most) DNA differences may not represent mutations at all; they may have been supernaturally created in Adam and Eve from the start — e.g., Adam and Eve would have been created with genetic differences. Thus, when we’re evaluating the billions of DNA letters in our cells and trying to determine when the differences began arising, it’s as if we were asked how long a clock has been ticking — but then were told that the clock has at least four hands instead of two.61
Therefore, to use DNA as a clock to measure when humanity began requires a very careful accounting of all potential means of genetic change and all potential genetic starting points. In other words, the only relevant DNA clock to the human origins debate is one in which evolutionists and creationists agree on the mechanism by which DNA differences arise as well as on the number of starting points from which DNA differences can arise.
Only one candidate DNA clock currently fulfills these criteria. Again, the vast majority of the billions of DNA letters in our cells do not lend themselves to a head-to-head comparison. Both sides may claim that the data fits their view, but claiming that the data support a view to the exclusion of the other is very challenging (as illustrated in the previous section).
Conversely, creationists and evolutionists agree on the origin of DNA differences in a tiny subsection of DNA (~16,559 DNA letters long) contained in the energy factories of our cells, called the mitochondria. Mitochondria and mitochondrial DNA (“mtDNA”) are found in both males and females, but only females appear to pass on mtDNA to their offspring. In other words, we each received our mtDNA from our mother, and our spouses received theirs from their mothers. Each of our children in turn did not inherit their father’s mtDNA; they inherited their mother’s.
Evolutionists agree that the current mtDNA differences among modern humans are traceable to a single woman in the past, whom they label “Eve.”62 However, they insist that this woman was part of a population of humans, not a single pair. This conclusion arises not because of anything inherent to the mtDNA data but because of the data from the billions of letters in the rest of the DNA sequence and the evolutionary presuppositions that we discussed in the previous section.
From a biblical perspective, all humans trace their ancestry back to Adam and Eve. However, because mtDNA is maternally inherited, YE creationists would agree with evolutionists that mtDNA differences today are traceable to a single woman in the past — Eve (both creation and evolution refer to her with the same name). Furthermore, both evolutionists and creationists would agree that modern mtDNA differences are the result of copying errors (i.e., mutations). Unlike the 3 billion DNA letters of DNA in the cell’s nucleus that come in two versions, mtDNA comes in only one version — effectively, the mother’s version. Hence, mtDNA differences arise via copying errors and were not created in Eve.
Thus, on the question of the origin of mtDNA differences, evolutionists and creationists are in complete agreement, except for one point — when this maternal ancestor lived. (Again, the evolutionary claims about this woman being part of a population have nothing to do with the mtDNA data itself; the population claim is imposed from the outside on top of the mtDNA data.)
To summarize up to this point, when we’re discussing mtDNA, both origins views hold to a single starting point. Because mtDNA comes in one version, not in two versions like the 3 billion letters of nuclear DNA sequence, both origins views also hold to copying errors (mutations) as the sole source of DNA variety (i.e., YE creationists do not believe that God created different mtDNA versions in Eve). Thus, mtDNA comparisons are one of the few type-1 experiments that can actually be performed to answer the question of when humanity began, and since the rate at which mutations occur in mtDNA has already been measured, this experiment can be performed right now.
To use mtDNA as a clock, we simply use this measured mutation rate to make testable predictions based on either the evolutionary timescale or on the YEC timescale and then compare the predictions with the scientific, observed facts. In other words, rather than starting with mtDNA differences in the present and then dialing the clock backward to see how long it would take to get to Eve, we’re going to go backward in time to the beginning under each model and predict what would have happened if the clock were allowed to run forward to the present. Specifically, we will assume for sake of argument that humans originated a long time ago (180,000 years ago under the evolutionary model63) or recently (4,500 years ago under the YEC model,64 representing the end of the Flood — see technical references for technical genetic reasons why the Flood date rather than the creation date was chosen).65 Then we will predict how many mtDNA differences should have accumulated in the timeframe specific to each model, after which we’ll compare these predictions to the actual number of differences in the current human population.
Thus, by multiplying the measured mutation rate of mtDNA66 by 180,000 years or by 4,500 years, we can make testable predictions about the timescale of human origins. Comparing these predictions to actual mtDNA differences at the global scale reveals a result that strongly contradicts the evolutionary timescale and confirms the YEC timescale (Figure 2).67
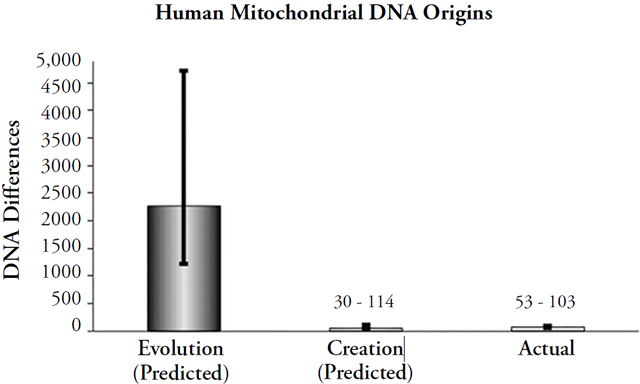
Figure 2. Comparison of origins predictions to actual human mitochondrial DNA differences. Differences were predicted by multiplying the measured mitochondrial DNA mutation rate by 2 and by the model-specific time of origin (e.g., for evolution, 180,000 years was used; for creation, 4,364 years was used as the (post-Flood) time of origin). The height of each column represents the average number of differences that would have accumulated under the model- specific time of origin (“Evolution” and “Creation”), and the black lines spanning the top of each column represent the full statistical range of each prediction, not the standard deviation (e.g., the lines represent the maximum best possible guesses under the evolutionary or creation timeframes). The height of the “Actual” column represents the average DNA differences in Africans today, and the black line spanning it represents the standard deviation. African DNA differences were used instead of non-African differences because Africans are the most genetically diverse group alive today and because evolutionists posit that Africans evolved first.
After 180,000 years, humans would have accumulated over 2,000 DNA differences (range = 1,220 to ~4,700)68 via the process of mutation to mtDNA. In just 4,364 years,69 humans would have accumulated only 30 to 114 mutations.70 Currently, about 78 differences exist on average in African populations (i.e., the most genetically diverse of all the human ethnic groups), with a maximum difference of ~120. Clearly, the YEC timescale accurately predicts the number of DNA differences that we observe today, while the evolutionary timescale predicts numbers an order of magnitude higher. Similar results hold true in animal species, as illustrated in Figure 3.
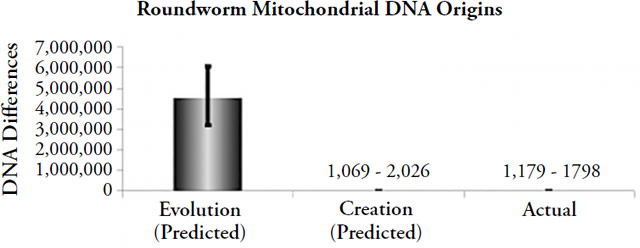
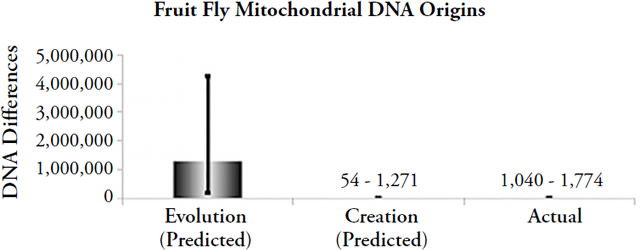
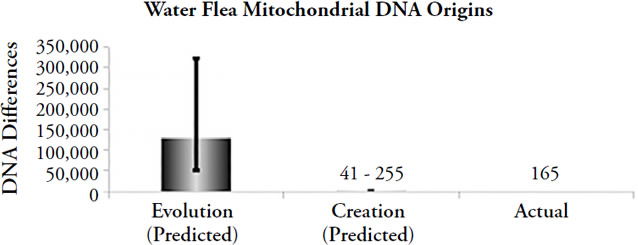
Figure 3. Comparison of origins predictions to actual animal mitochondrial DNA differences. Differences were predicted by multiplying the measured mitochondrial DNA mutation rate by 2 (roundworms, fruit flies) or by 1 (water fleas), and by the model-specific time of origin (e.g., for evolution, the time appropriate to each organism was used; for creation, 6,000 years was used as the time of origin). The height of each column represents the average number of differences that would have accumulated under the model-specific time of origin (“Evolution” and “Creation”), and the black lines spanning the top of each column represent the full statistical range of each prediction, not the standard deviation (e.g., the lines represent the maximum best possible guesses under the evolutionary or creation timeframes). The height of the “Actual” column represents the average DNA differences today, and the black line spanning it represents the range of differences (where appropriate).
These findings represent much more than an isolated, irrelevant data point in the bigger creation/evolution debate. As we observed above, mtDNA is one of the only arenas in which a straightforward type-1 experiment can be performed — one of the only arenas in which we can judge the scientific validity of the creation model versus the evolution model. Furthermore, performing this mtDNA experiment in a wide variety of animal species leads to the same conclusion: the biblical view of earth history is correct.71 Thus, the evolutionary timescale runs into trouble not only on the question of human origins but across a much wider swath of biological life.
Implicit in these calculations was the assumption that the mtDNA mutation rate has been constant with time. We made this assumption since it forms the basis for the entire millions-of-years paradigm in the evolutionary model. When evolutionists claim that the earth or the universe is ancient, their methods assume that the geologic or astronomical processes that they observe today have occurred at a constant rate throughout the history of the earth or universe.72
For decades, YE creationists have pointed out the arbitrary nature of this assumption,73 especially in light of the global Flood element of the YEC model of geology.74 Essentially, YE creationists have correctly identified the entire millions-of-years paradigm as nothing more than a type-3 experiment. In short, the evolutionary argument about the age of the earth and of the universe work only if the assumption about constant rates of change is true. Change that assumption and the entire paradigm collapses.
Thus, by assuming constant rates of genetic change in our calculations, we made the calculations overly generous to the evolutionary view. The fact that the evolutionary predictions could not be reconciled with reality even under generous assumptions makes the explanatory dilemma for evolutionists all the greater. If they claim that rates of genetic change were different in the past, they’ve just undermined the foundational assumption of their entire ancient universe/ancient earth view. If they do nothing, they are left with a glaring contradiction between predictions and facts. Hence, these mtDNA results have implications for the evolutionary view far beyond biology, and they make the evolutionary paradigm even harder to maintain in a scientifically consistent and coherent way.
Perhaps the evolutionists will invoke natural selection to explain why their predictions do not match up with facts. In other words, perhaps humans have fewer genetic differences than predicted under the evolutionary model because natural selection eliminated a number of copying errors that arose in the past. This hypothesis would be worth exploring — but only if it leads to testable, falsifiable predictions.
Summary
In summary, there is no genetic evidence to support an ancient origin for mankind. The DNA differences in the billions of DNA letters in the cellular compartment termed the nucleus are easily explicable from two people in the last 6,000 years (see previous section), and the mtDNA differences observable today are all the more explicable (Table 4; Figure 2). The mtDNA arena of comparison also happens to be one arena in which a type-1 experiment can be performed, and the evidence strongly contradicts the evolutionary timescale while confirming the YEC timescale. Since these results assumed constant rates of genetic change, and since evolutionary geology and astronomy also depend on the assumption of constant rates of change for their millions- and billions-of-years conclusions, these genetic findings throw into confusion these two fields of physical science as well. Genetically speaking, mankind appears to have originated only a few thousand years ago.
| Cellular compartment | Letters in DNA sequence | Inheritance | Origin of human-human differences under YEC view |
|---|---|---|---|
| Nucleus | 3,000,000,000 | Paternal and Maternal | Majority of DNA differences due to Creation, minority due to mutation |
| Mitochondria | 16,559 | Maternal | All DNA differences due to mutation |
Again, the success of these initial genetic results gives us confidence that we can predict mtDNA mutation rates for other species, and we are willing to test these predictions in the lab. In fact, we invite our evolutionary colleagues to join us so that we can perform a type-1 experiment as accurately as possible. If our evolutionary colleagues are unwilling or unable to make and test a falsifiable prediction, why should we view their claims as scientific rather than pseudoscientific?
IV. Where: Africa or Ararat?
The mtDNA results discussed above hinted at the one element of human origins that we have not explored in detail — the timing and geography of the origin of African people groups. On the question of geography, creation and evolution are largely in agreement — except for the origin of African people groups. Evolutionists posit that Africans evolved first and then gave rise to the non-African groups.75 In contrast, YE creationists posit the simultaneous origin of the major ethnic groups very soon after the dispersion at the Tower of Babel.
The genetic aspects of the evolutionary claim rests on a technical aspect of mtDNA comparisons. Both evolutionists and creationists use software to visualize the number of DNA differences among various individuals or ethnic groups, and one of the most common visualization tools is the creation of phylogenetic or family trees. Naturally, this implies a genealogical relationship among those connected on the tree, but, in the software employed, ancestry assumptions are not necessary. The tree simply depicts the number of DNA differences in a visually striking way.
When the evolutionists draw trees, they of course assume common ancestry regardless of the species compared, since one of the foundational tenets of evolution is universal common ancestry of all species on earth (i.e., all plants, animals, and humans are descended from a single common and microscopic ancestor). Not surprisingly, when evolutionists draw family trees of the human ethnic groups using mtDNA comparisons, they include chimpanzee DNA.76 This resultant tree — which evolutionists interpret as genealogical relationships — shows some of the African branches splitting off first (about 120,000–180,000 years ago, as we alluded to in section III) followed by non-African groups later (about 50,000 years ago).
Even if you omit the chimpanzee DNA from the comparison and draw the tree using only modern human ethnic groups, it is still obvious that African ethnic groups have about twice as many mtDNA differences among them as do non-African ethnic groups. If you assume that the rate of mtDNA mutations is constant with time, the fact of greater mtDNA diversity in Africans implies that Africans have been around longer than non-Africans.
However, implicit in this conclusion is a technical assumption about the mtDNA mutation rates. To measure these rates empirically, scientists must use pedigrees,77 which means that the units are reported in terms of mutations per generation. To convert these units to absolute time (i.e., mutations per year), scientists must make an assumption about how many years pass per generation. Evolutionists implicitly assume that the generation times (time from birth of parent to birth of child) across all ethnic groups are the same.
However, marriage data from the United Nations suggests that this assumption is not valid (Table 5).78 On average, African females marry earlier in life than non-African females. About 32% of African women are married by ages 15–19, whereas only 12% of non-African women are married by the same age. This roughly three-fold difference disappears at later ages (e.g., about the same number of African and non-African women are married by their 30’s and 40’s), suggesting that the generation time in Africans might be about twice as fast as the generation time in non-Africans. Since mtDNA is passed on maternally, these data imply that some African ethnic groups have twice as many mtDNA differences because twice as many generations have passed in their lineages as compared to non-African lineages.
| Age Bracket | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 15–19 | 20–24 | 25–29 | 30–34 | 35–39 | 40–44 | 45–49 | 50–54 | 55–59 | 60–64 | 65+ | |
| Africa: % of women married | 32.0 | 67.4 | 81.3 | 83.3 | 83.6 | 79.2 | 74.4 | 64.7 | 56.8 | 44.4 | 28.2 |
| non-Africa: % of women married | 11.8 | 47.2 | 69.8 | 77.0 | 78.2 | 77.0 | 73.8 | 67.7 | 61.6 | 51.0 | 32.2 |
| Fold-difference | 2.7 | 1.4 | 1.2 | 1.1 | 1.1 | 1.0 | 1.0 | 1.0 | 0.9 | 0.9 | 0.9 |
The data we presented in figure 2 made predictions for a variety of generation times (e.g., 15 years to 35 years). Under none of these generation times could the evolutionary model correctly predict the amount of DNA differences observable today. In contrast, the YEC predictions correctly predicted the African mtDNA differences under the assumption of a higher generation time (e.g., assuming a generation time of 15 years, the YEC model predicts 69 to 114 DNA differences in 4,364 years, which captures the average mtDNA differences — 78 — present today among Africans). The mtDNA differences among non-Africans (about 49, not displayed in figure 2) were predictable under the YEC model by assuming a generation time of 25 years (predicted range of differences = 41 to 69). Thus, the fact of higher mtDNA diversity in Africans does indeed appear to be due to their earlier age of marriage (and, presumably, of child-bearing), not to their supposed ancient evolutionary origin.
These data notwithstanding, evolutionists have also tried to buttress their out-of-Africa claims with data from the 3 billion DNA letters in the genome of the cell nucleus that we discussed previously — the main engine of heritability and diversity among humans. Specifically, Africans have more DNA differences among these 3 billion letters than non-Africans (only about 1.25-fold more), and they have more combinations of these differences (in technical genetic terms, linkage disequilibrium is lower in Africans).79 To the evolutionist, these facts are consistent with an ancient origin of humans in Africa, and a more recent population bottleneck in their descendants who left Africa to found the modern non-African ethnic groups.
Again, these claims rest on assumptions of identical generation times among African and non-African ethnic groups, an assumption that is not borne out by current data. In addition, it appears that Africans reshuffle (e.g., in technical terms, recombine) their DNA at higher rates and/or in different places than non-Africans, which would explain their extra combinations (e.g., lower linkage disequilibrium) of DNA — a conclusion that even the evolutionary community concedes.80
About the only genetic arena in which evolutionists can still hope to find evidence for an early origin of mankind out of Africa is in the Y chromosome — the chromosome unique to males, which is passed from fathers to sons. Current data indicate that African men have about twice as many Y chromosome differences as non-African men.81 However, the rate at which the Y chromosome changes — either by mutation or by a process termed gene conversion — has not been published for Africans. We predict that African Y chromosomes will change twice as fast as non-African Y chromosomes. Conversely, if evolutionists are confident in their out-of-Africa model of human origins, we invite them to make a counter-prediction — and then test their ideas with us in the lab.
In summary, there is no straightforward genetic evidence for the origin of mankind first in Africa. Evolutionists reach this conclusion genetically by assuming human-ape common ancestry and by assuming that the generation times of all ethnic groups are identical. In the context of the origins debate, the first assumption represents circular reasoning, and the second assumption does not match published data. Africans reproduce earlier than non-Africans and reshuffle their DNA faster/in more places than non-Africans, and both of these facts appear sufficient to explain the data that we observe without invoking separate times of origin for the various people groups in existence today (Table 6).
| Cellular compartment | Inheritance | Genetic differences between Africans and non-Africans | Facts demonstrating contemporaneous origin of African and non- African people groups | Prediction |
|---|---|---|---|---|
| Nucleus | Paternal and Maternal | 1.25-fold | Africans reshuffle their DNA faster (promotes retention of DNA differences) | |
| Mitochondria | Almost exclusively Maternal | 1.5- to 2-fold | As compared to non-African women, twice as many African women marry early (more generations have passed in Africans, leading to more DNA differences) | |
| Y chromosome | Paternal | 2-fold | Y chromosomes in Africans mutate/undergo gene conversion faster than in non-Africans |
Why Don’t More Scientists Accept These Conclusions?
The conclusions that we’ve presented in this chapter are obviously at odds with the dominant scientific paradigm in the Western world today. How can our claims possibly be true? Evolutionists have an explanation that they’ve advanced for decades: YEC conclusions are not true. The justification that evolutionists cite for this claim is the absence of YEC conclusions from the mainstream peer-reviewed scientific literature. And why are creationist conclusions absent from this literature? The quote from BioLogos that we cited above is worth repeating here:
The reason Christian anti-evolutionary approaches are absent from the mainstream scientific literature is not because scientists are theologically or philosophically biased against them, but rather because they offer little in the way of useful tools for making accurate predictions about the natural world.82 [emphasis added]
As we’ve observed, this is factually untrue. In the realm of science that we’ve briefly examined in this chapter, YE creationists make many testable, accurate predictions about the natural world, and it’s the evolutionists who historically have had trouble getting their predictions to match facts.
Furthermore, YE creation scientists do not publish un-reviewed technical papers. The major scientific players in the YEC field all earned their degrees from reputable secular universities with many also having many secular publications prior to making a career shift into origins research,83 and we submit our findings to one another for peer-review prior to publication. Just like the secular peer-review system, some of our initial conclusions must be significantly refined or rejected before they have a chance of being published.
Naturally, evolutionists might criticize YEC scientists relying on like-minded individuals (e.g., fellow YEC scientists) for the peer-review process. Evolutionists might claim that this represents a self-reinforcing process that is ultimately flawed and useless to scientific progress. But YEC scientists could say the same about evolutionists. The latter do not consult with YEC scientists before publishing their evolutionary conclusions. Instead, they solicit the assistance and review of the fellow, like-minded evolutionists!
Thus, on two counts, the common evolutionary reason for the absence of creationist ideas from mainstream scientific literature is wrong. First, creationists do indeed submit their research to peer review. Second, as we have demonstrated, they make testable scientific predictions that, in many cases, are more accurate than the predictions of the evolutionists (e.g., see preceding sections).
The latter fact raises an important question: Why don’t evolutionists submit their ideas to creationist peer-review before publication? Why not solicit YEC PhD scientists for help and criticism before publishing a paper? Why not consult with the YEC community (at least informally) before taking evolutionary ideas public? Doing so might save the evolutionary model from further erroneous predictions.
To answer the question that heads this section, the BioLogos claim that we cited above would suggest that we are left with only one option: The vast majority of professional scientists are theologically or philosophically biased against creationist ideas. At first pass, this would seem conspiratorial and, therefore, difficult to accept.
Yet upon further reflection, this wooden interpretation of our options becomes much more nuanced in light of a few key facts. First, surveys show that the vast majority of scientists are unbelievers. Nearly 70% of scientific professionals cannot positively say that they believe in God.84 Since belief in God is a necessary (but insufficient) profession for one to be a Christian, the number of non-Christian scientists is likely even higher than 70%.
Second, Scripture tells us that unbelievers do indeed have a bias. “For the wrath of God is revealed from heaven against all ungodliness and unrighteousness of men, who suppress the truth in unrighteousness, because what may be known of God is manifest in them, for God has shown it to them. For since the creation of the world His invisible attributes are clearly seen, being understood by the things that are made, even His eternal power and Godhead, so that they are without excuse” (Rom 1:18–20). Not only do unbelievers suppress the truth about God, they suppress the truth about God that is revealed in nature. Thus, the creation/evolution debate is at the heart of the unbeliever’s dealings with God.
However, this passage in Romans does not suggest that all unbelievers go on the warpath against creationist ideas. Instead, Scripture says that unbelievers suppress the truth; they don’t all violently try to destroy it. Conversely, suppressing the truth can take many forms — from passively ignoring contrary ideas, to never attempting to learn or understand uncomfortable contrary claims, to occasionally expressing strong dislike for an idea. In other words, non-Christian scientists are much like the unbelievers that we encounter every day. Most are passively disinterested in and ignorant of the things of God and of the scientific ramifications of the creation account. Only a few are visibly and adamantly opposed.
Sadly, as the above discussion demonstrates, evolutionists who are professing Christians appear to practice the same behavior.85 For example, they seem to never have considered alternative hypotheses on the question of ancestral population size, and they regularly and prematurely turn highly speculative hypotheses into fact (e.g., Table 2).
The latter error should technically be termed pseudoscience. However, since the evolutionary creationists we cited are well trained and practiced scientists, we don’t think this error stems from any lack of quality training. Instead, it is more likely to stem from an ignorance of the opposition. In other words, when a scientist is completely unaware of a contrary view, his hypothesis may seem like fact since nothing else seems able to explain the data he’s observing.
In support of this conjecture, mainstream evolutionary literature demonstrates ignorance of creationist ideas.86 For example, evolutionists regularly contend that accepting YEC requires throwing out science entirely:
If someone challenges the current paradigm of [sic] by asserting that the Earth is not 4.5 billion years old, but rather was created by divine intervention 6000 years ago . . . the correct response is: “Well, maybe. But if that is what happened, then much else of what we think we know must also be wrong. We will need a new explanation for how the Sun gets its energy, as our laws about nuclear physics must be wrong. As this is the physics that has manifestly empowered engineers to build nuclear power plants, we need to explain how they are doing so well even though they are operating with the incorrect laws. The same would go for the empowerment provided by science for the use of radioisotopes in medicine X-rays in dentistry.87
The former president of BioLogos repeats this claim:
The conclusion that creation is ancient does not come from interpretations at the periphery of these disciplines; it is at the core of all that nuclear physicists, geologists and astronomers do every day. For you or I to say that they are wrong is to say that these entire disciplines — geology, nuclear physics and astronomy — have got almost everything wrong.88
But it is nearly impossible to read and understand the YEC scientific literature and arrive at the conclusions above. These claims — that accepting YEC requires throwing out physics, geology, etc. — are as far from the truth as any stereotype of YEC science can be. Since we are confident that both of the men responsible for the quotes above are scholarly and logical scientists, we are left with one option: they haven’t read and/or understood the YEC scientific literature.
Even more disappointing, the few evolutionary creation scientists with whom the authors of this chapter have personally communicated seem to have no interest in the YEC scientific literature. When we’ve presented them with the opportunity to engage the scientific data (e.g., by pleading with them to rigorously peer-review creationist findings before publication), they have declined. One theistic evolutionist has even admitted a past bias toward opponents, confessing that he viewed them as dumb and uninformed. If this is how professing Christians behave when confronted with contrary evidence, how much more so the unbelieving scientists!
In sum, the vast majority of the scientific world is at odds with the conclusions that we have presented here about human genetic origins because they appear to never have educated themselves on their opponents’ scientific positions. More troubling is that, in some cases, evolutionists appear to have even deliberately avoided the opposition, and in the most extreme cases, intentionally suppressed it.89 While this phenomenon could be labeled “bias,” it does not appear to involve a deliberate and planned conspiracy among scientists in the Western world. Instead, for unbelievers, it appears to flow from their deeply rooted spiritual state. Since unbelievers are too proud to acknowledge God in their thinking,90 and since all believers, ourselves included, are in the process of sanctification and can fall prey to some of the same sins that unbelievers practice, such as spiritual and/or intellectual pride,91 the fear of man, and the desire for academic respect from the secular world,92 “pride” rather than “bias” may be the better answer to the question that heads this section.
Summary and Ramifications
From the brief overview of the technical scientific literature that we’ve sketched, three facts emerge. First, the evolutionary model of human origins has a long history of scientific failure (Tables 2–3). It has repeatedly made public pronouncements of fact only to discover new data that contradict these claims. Hence, before we can even explore the question of whether evolution works as a scientific model today, we are struck with the dismal track record of evolution in times past.
Second, the evolutionary model does a poor job of explaining data in the present (Figure 2). When pressed to explain human-human genetic differences observable today, evolutionary predictions are an order of magnitude off the actual value. In essence, the evolutionary model cannot predict the rate of mtDNA mutation in humans. Since mutations are supposed to be the engine of evolution and the driver of all evolutionary change, this mismatch between predictions and facts is all the more profound.
Third, the YEC conclusions that we’ve highlighted in this chapter represent a comprehensive answer to the question of human genetic origins. Our claims and observations encompass virtually every genetic compartment present in human cells (Table 7), and they account for the millions of DNA differences across ethnic groups present in the world today. Furthermore, they robustly answer the questions of from whom humans originated (people, not apes), how many humans began our species (two — Adam and Eve), when humans originated (about 6,000 years ago), and where major human ethnic groups originated (near Ararat). In short, they explain all the data for which we have experimental results. For those areas in which experiments are forthcoming, we presented testable predictions that can be falsified in the lab (e.g., Table 5).
| Type of Genetic Comparison | YEC Status |
|---|---|
| Human vs. human nuclear DNA | Successful prediction of mutation, genetic reshuffling rate (e.g., recombination & gene conversion) for entire sequence |
| Human vs. human mitochondrial DNA | Successful prediction of mutation rate |
| Human vs. human Y chromosome | Pending prediction for Y chromosome mutation/genetic reshuffling (e.g., gene conversion) rate |
In light of these facts, it is all the more remarkable that evolutionists can continue to accuse creationists of being ignorant of the “big picture” of evolution. While this chapter covers only the question of human genetic origins, the accompanying chapters demonstrate the veracity of the biblical account of human origins from a variety of fields. To say that creationists are only capable of finding minor holes in evolutionary arguments while missing the larger body of evidence is unjustifiable.
Furthermore, the claim that “multiple independent lines of genetic evidence” support human evolution is false. Again, evolutionists are fond of appealing to the “big picture” when confronted with a contradiction between one of their predictions and fact. Logically, if every one of their claimed evidences fails, then the sum of these broken evidences cannot possibly add up to a successful model. As we have observed, all the claimed evolutionary evidences represent type-3 experiments, or they represent type-2 experiments that could falsify or have already falsified evolutionary predictions (rather than YEC predictions) (e.g., Tables 2–4, 6). Multiple independent lines of evidence demonstrate that evolutionary claims are unscientific.
As described at the beginning of this chapter, the gold standard of science is the ability of a model to make testable accurate predictions. From the assumptions of the YEC model, creationists have made testable predictions about the future that can be tested in the laboratory. If evolutionists have a problem with what we’ve concluded, we’ve given them a ready means by which to falsify our position. In other words, the YEC model of genetics has matured into a full-fledged scientific alternative to the evolutionary model, with much stronger predictive power.
Furthermore, the conclusions in this chapter represent only a fraction of the mature YEC model. We’re in the process of publishing testable genetic predictions for a great assortment of animal species alive today for which genetic data is available.93 The “big picture” of evolution can now be compared head-to-head with the “big picture” of YEC — if evolutionists are able to come up with some falsifiable predictions of their own.
In light of these advances, we would be fully justified in taking the evolutionists’ criticisms of creation right back to them. If evolutionists want to be taken seriously in the origins debate, then they need to do more than make an isolated claim about an obscure species here and there that shows nothing but shifts in existing genetic variation or an isolated benefit due to the loss of genetic information. Instead, they need to give us a comprehensive model, a falsifiable explanation that accounts for the genetics of all species alive today. Science demands no less.
Searching for Adam

You can believe what the Bible says about Adam and man's origin and with intellectual integrity reject the myth of human evolution.
Read Online Buy Book
Master Books has graciously granted AiG permission to publish selected chapters of this book online. To purchase a copy please visit our online store.
Footnotes
- Nathaniel Jeanson received a B.S. in Molecular Biology and Bioinformatics from the University of Wisconsin-Parkside and Ph.D. in Cell and Developmental Biology from Harvard University. Jeff Tomkins received his B.S. in Agricultural Education from Washington State Univ., an M.S. in Plant Science from Univ. Idaho, and a Ph.D. in Genetics from Clemson University.
- Michael Buratovich, “Biological Evolution: What Makes it Good Science? Part 1,” https://BioLogos.org/blogs/archive/biological-evolution-what-makes-it-good-science-part-1. See also chapter 23 of Douglas J. Futuyma, Evolution (Sunderland, MA: Sinauer Associates, Inc., 2013). “The most important feature of scientific hypotheses is that they are testable” (emphasis his, p. 635).
- Dennis Venema, “Theory, Prediction and Converging Lines of Evidence, Part 3.” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/theory-prediction-and-converging-lines-of-evidence-part-3.
- The Chimpanzee Sequencing and Analysis Consortium, “Initial Sequence of the Chimpanzee Genome and Comparison with the Human Genome,” Nature 437 (2005): 69–87, http://www.nature.com/nature/journal/v437/n7055/full/nature04072.html.
- Kevin E. Langergraber et al., “Generation Times in Wild Chimpanzees and Gorillas Suggest Earlier Divergence Times in Great Ape and Human Evolution,” Proceedings of the National Academy of Sciences USA 109, no. 39 (2012): 15716–15721, http://www.pnas.org/content/109/39/15716.full; Oliver Venn et al., “Strong Male Bias Drives Germline Mutation in Chimpanzees,” Science 344 (2014): 1272–1275.
- Eve was “the mother of all living” (Gen. 3:20) and from the eight people on the ark “the whole earth was populated” (Gen. 9:19).
- As is shown in the later chapter by Bergman, biblically speaking there is only one race, Adam’s race.
- Note that the genealogies of Shem, Ham, and Japheth in Genesis 10 abruptly end after a few generations — consistent with the writer of Genesis 10 being unable to communicate with the members of additional generations due to a language barrier brought about by the Tower of Babel incident.
- Chris Hardy and Robert Carter, “The Biblical Minimum and Maximum Age of the Earth,” Journal of Creation 28, no. 2 (2014): 89–96, http://creation.com/images/pdfs/tj/j28_2/j28_2_89-96.pdf; Robert Carter and Chris Hardy, “Modelling Biblical Human Population Growth,” Journal of Creation 29, no. 1 (2015): 72–79. Since Peleg was born 101 years after the flood and lived 209 years, and we are told that the division of humanity at the Tower of Babel was some unspecified date “in the days of ” Peleg, we cannot be precise on the dating of the division.
- John R. Grehan and Jeffrey H. Schwartz, “Evolution of the Second Orangutan: Phylogeny and Biogeography of Hominid Origins,” Journal of Biogeography 36 (2009): 1823–1844.
- DNA repair machinery exists in the cell, but some copying mistakes still apparently slip through each generation.
- For the chimpanzee reference, see Oliver Venn et al., “Strong Male Bias Drives Germline Mutation in Chimpanzees,” Science 344 (2014): 1272–1275. The human rate has been measured on multiple occasions; for an example, see Donald F. Conrad et al., “Variation in Genome-wide Mutation Rates Within and Between Human Families,” Nature Genetics 43, no. 7 (2011): 712–714, http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3322360/.
- An example of the math: 3,000,000 years / 20 years per generation = 150,000 generations.
- An example of the math used to derive the figure of “80 million DNA letters apart” is as follows. In 13,000,000 years, about 650,000 generations pass [13,000,000 years / 20 years per generation = 650,000 generations]. Using this number, 60 DNA changes per human generation x 650,000 generations = 39,000,000 DNA changes total. Since the identical process would occur in chimpanzees, the total would need to be multiplied by two [39,000,000 x 2 = 78,000,000 DNA changes total in both the human lineage and the chimpanzee lineage]. Rounding numbers, the total is ~80,000,000 DNA changes in 13,000,000 years.
- Technically, since our DNA comes in two versions (in technical terms, we are a dipoid species — versus a haploid species), humans have 6 billion total DNA letters (as do chimps). But 3 billion is a useful simplification for our purposes in this section.
- An example of the math used: 78,000,000 predicted DNA differences between humans and chimpanzees / 3,000,000,000 total DNA letters in humans = 0.026 = 2.6%, or about 3% difference.
- Jeffrey P. Tomkins, “Genome-Wide DNA Alignment Similarity (Identity) for 40,000 Chimpanzee DNA Sequences Queried against the Human Genome is 86–89%,” Answers Research Journal 4 (2011): 233–241, https://answersingenesis.org/genetics/dna-similarities/genome-wide-dna-alignment-similarity-identity-for-40000-chimpanzees/; Jeffrey P. Tomkins, “Documented Anomaly in Recent Versions of the BLASTN Algorithm and a Complete Reanalysis of Chimpanzee and Human Genome-Wide DNA Similarity Using Nucmer and LASTZ,” Answers Research Journal 8 (2015): 379–390, https://answersingenesis.org/genetics/dna-similarities/blastn-algorithm-anomaly/.
- In the 2005 Nature paper describing the elucidation of the chimpanzee DNA sequence (accessible at http://www.nature.com/nature/journal/v437/n7055/full/nature04072.html), the authors stated, “Best reciprocal nucleotide-level alignments of the chimpanzee and human genomes cover ~2.4 gigabases (Gb) [2,400,000,000 DNA letters] of high-quality sequence, including 89 Mb [89,000,000 DNA letters] from chromosome X and 7.5 Mb [7,500,000 DNA letters] from chromosome Y” (p.71). Only these 2,400,000,000 DNA letters were used to calculate the published 1.23% DNA difference between humans and chimpanzees. In table 1 of the same paper, it is clear that 2.7 gigabases (GB) — 2,700,000,000 DNA letters — in total were sequenced, leaving 0.3 GB — 300,000,000 DNA letters (about 10% of 3 billion) — unaccounted for, consistent with Jeff Tomkins’ independent findings. Furthermore, by last count (http://www.ncbi.nlm.nih.gov/genome/, accessed 09/28/15), the total number of DNA letters in chimpanzees is 3,309,000,000, and in humans it is 3,259,520,000 DNA letters, leaving even more potential DNA differences unaddressed. Clearly, a DNA difference between humans and chimpanzees of 1.23% represents a careful selection of a subset of the facts.
- Assuming that humans possess 3,259,520,000 total DNA letters, a 12% DNA difference from apes (the result of only 88% identity — see previous footnotes) entails the following: 0.12 x 3,259,520,000 total DNA letters = 391,142,400 DNA letters difference, which is about 400 million DNA differences between chimps and humans.
- Kevin E. Langergraber et al., “Generation Times in Wild Chimpanzees and Gorillas Suggest Earlier Divergence Times in Great Ape and Human Evolution,” Proceedings of the National Academy of Sciences USA 109, no. 39 (2012): 15716–15721, http://www.pnas.org/content/109/39/15716.full.
- Oliver Venn et al., “Strong Male Bias Drives Germline Mutation in Chimpanzees,” Science 344 (2014): 1272–1275. We give special thanks to Rob Carter for bringing this evolutionary discrepancy to our attention.
- For example, see Michael Buratovich, “Biological Evolution: What Makes it Good Science? Part 1,” https://BioLogos.org/blogs/archive/biological-evolution-what-makes-it-good-science-part-1; for a non-BioLogos reference see Douglas J. Futuyma, Evolution (Sunderland, MA: Sinauer Associates, Inc., 2013).
- Nathaniel T. Jeanson, “Darwin vs. Genetics: Surprises and Snags in the Science of Common Ancestry,” Acts & Facts 43, no. 9 (2014): 8–11, http://www.icr.org/article/darwin-vs-genetics-surprises-snags.
- For example, “Humans share more DNA with chimpanzees than with any other animal, suggesting that humans and chimps share a relatively recent common ancestor.” See Anon., “What Is the Genetic Evidence for Human Evolution?” https://BioLogos.org/common-questions/human-origins/what-scientific-evidence-do-we-have-about-the-first-humans/.
- See Anon., “Genetics,” http://biologos.org/resources/audio-visual/genetics, and Dennis Venema, “Theory, Prediction and Converging Lines of Evidence, Part 3,” http://biologos.org/blogs/dennis-venema-letters-to-the-duchess/theory-prediction-and-converging-lines-of-evidence-part-3.
- Diagrams for this supposed evolutionary event typically show the fusion of only one member of each pair.
- Jerry Bergman and Jeffrey Tomkins, “The Chromosome 2 Fusion Model of Human Evolution — Part 1: Re-evaluating the Evidence,” Journal of Creation 25, no. 2 (2011): 106–110, http://creation.com/chromosome-2-fusion-1; Jeffrey Tomkins and Jerry Bergman, “The Chromosome 2 Fusion Model of Human Evolution — Part 2: Re-analysis of the Genomic Data,” Journal of Creation 25, no. 2 (2011): 111–117, http://creation.com/chromosome-2-fusion-2; Jeffrey Tomkins, “Alleged Human Chromosome 2 ‘Fusion Site’ Encodes an Active DNA Binding Domain Inside a Complex and Highly Expressed Gene — Negating Fusion,” Answers Research Journal 6 (2013): 367–375, https://answersingenesis.org/genetics/dna-similarities/alleged-human-chromosome-2-fusion-site-encodes-an-active-dna-binding-domain-inside-a-complex-and-hig/.
- Dennis Venema, “Signature in the Synteny,” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/signature-in-the-synteny.
- Michael D. Wilson et al., “Species-specific Transcription in Mice Carrying Human Chromosome 21,” Science 322 no. 5900 (2008): 434–438, http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3717767/; Nathaniel T. Jeanson, “An Update on Chromosome 2 ‘Fusion,’ ” Acts & Facts 42 no. 9 (2013): 13, http://www.icr.org/article/update-chromosome-2-fusion.
- Anon., “What Is the Genetic Evidence for Human Evolution?” https://BioLogos.org/common-questions/human-origins/what-scientific-evidence-do-we-have-about-the-first-humans/. Evidence #2 references “genetic scars” and implicitly assumes that these sorts of genetic differences (the “scars”) represent mutated and non-functional or functionally neutral sequences. For the “genetic synonyms” argument to work in Evidence #3, the argument must assume that these “synonyms” represent functionally neutral sequences.
- Nathaniel T. Jeanson, “Recent, Functionally Diverse Origin for Mitochondrial Genes from ~2700 Metazoan Species,” Answers Research Journal 6 (2013): 467–501, https://answersingenesis.org/genetics/mitochondrial-dna/recent-functionally-diverse-origin-for-mitochondrial-genes-from-~2700-metazoan-species/.
- BioLogos Editorial Team. “On Reading the Cell’s Signature,” https://BioLogos.org/blogs/archive/on-reading-the-cells-signature; Dennis Venema, “Understanding Evolution: Is There ‘Junk’ in Your Genome? Part 1,” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/understanding-evolution-is-there-junk-in-your-genome-part-1; Dennis Venema, “Is There ‘Junk’ in Your Genome? Part 2,” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/is-there-junk-in-your-genome-part-2.
- ENCODE Project Consortium, “Identification and Analysis of Functional Elements in 1% of the Human Genome by the ENCODE Pilot Project,” Nature 447 (2007): 799–816.
- ENCODE Project Consortium, “An Integrated Encyclopedia of DNA Elements in the Human Genome,” Nature 489 (2012): 57–74, http://www.nature.com/nature/journal/v489/n7414/full/nature11247.html).
- Ed Yong, “ENCODE: the Rough Guide to the Human Genome,” http://blogs.discovermagazine.com/notrocketscience/2012/09/05/encode-the-rough-guide-to-the-human-genome/#.V_AxDtx4yoI.
- For example, see Dan Graur et al., “On the Immortality of Television Sets: ‘Function’ in the Human Genome According to the Evolution-free Gospel of ENCODE,” Genome Biology and Evolution 5 no. 3 (2013): 578–590, http://gbe.oxfordjournals.org/content/5/3/578; for a response to Graur et al., see: Nathaniel Jeanson and Brian Thomas, “The Resurrection of ‘Junk DNA’?” http://www.icr.org/article/resurrection-junk-dna.
- Jeffrey P. Tomkins, “The Human Beta-Globin Pseudogene Is Non-Variable and Functional,” Answers Research Journal 6 (2013): 293–301, https://answersingenesis.org/genetics/human-genome/the-human-beta-globin-pseudogene-is-non-variable-and-functional/.
- Rachel Held Evans and Dennis Venema, “Ask an Evolutionary Creationist: A Q&A with Dennis Venema,” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/ask-an-evolutionary-creationist-a-qa-with-dennis-venema.
- Jeffrey P. Tomkins, “Challenging the BioLogos Claim that a Vitellogenin (Egg-Laying) Pseudogene Exists in the Human Genome,” Answers Research Journal 8 (2015): 403–411, https://answersingenesis.org/genetics/dna-similarities/challenging-BioLogos-claim-vitellogenin-pseudogene-exists-in-human-genome/.
- Ibid.
- In his multi-part response (http://biologos.org/blogs/dennis-venema-letters-to-the-duchess/series/vitellogenin-and-common-ancestry) to Tomkins’ rebuttal of the egg yolk gene claim, Venema tried to skirt Tomkins’ main point — that the actual molecular (e.g., DNA letter by DNA letter) evidence supporting the existence of an egg yolk gene remnant in humans is nonexistent. Tomkins noted that “Sequence identity [between human DNA and chicken DNA] dropped as the fragment size increased, eventually leveling off to about 39% identity.” In other words, when comparing the chicken egg yolk gene DNA sequence to human “egg yolk gene” DNA sequence, the match between the two is barely different from a random DNA match (25% identity represents a random DNA match). Even if we focus only on the few parts of the DNA where DNA sequence identity is higher, Tomkins’ noted that “Even in an evolutionary sense, to say that a pseudogene can be identified by only 0.35% of the original sequence is quite a stretch of the Darwinian paradigm.” In response to these data, Venema never provided any numbers to rebut Tomkins. Instead, Venema republished the diagrams from the original egg yolk gene paper, devoid of any percent identity labels. In other words, Tomkins reanalyzed the raw data and reported a serious criticism of the facts. In response, rather than deal with the facts, Venema created diagrams to make the similarity between chicken and human appear high — without actually constraining his depictions with numbers. At best, this is a tacit concession of defeat; at worst, it’s deliberately deceptive. In addition to diagrams devoid of numbers, Venema attempted to corral other lines of evidence to support his contention, but these “evidences” were simply reassertions of why he expected a broken egg yolk gene to exist in humans — an expectation that has been falsified by the evidence.
- Dennis Venema, “ENCODE and ‘Junk DNA,’ Part 1: All Good Concepts are Fuzzy,” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/encode-and-junk-dna-part-1-all-good-concepts-are-fuzzy; Dennis Venema, “ENCODE and ‘Junk DNA,’ Part 2: Function: What’s in a Word?” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/encode-and-junk-dna-part-2-function-whats-in-a-word.
- Dennis Venema, “Common Ancestry, Nested Hierarchies, and Parsimony,” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/adam-eve-and-human-population-genetics-part-6-common-ancestry-nested-hierarchies-and-parsimony.
- See Figure 4A of Cristina Sisu et al., “Comparative Analysis of Pseudogenes Across Three Phyla,” PNAS 111 (2014): 13361–13366, http://www.pnas.org/content/111/37/13361.long.
- Epigenetics is the study of heritable changes that do not involve changes in DNA sequence.
- E.g., a population living at the time that the evolutionists propose — hundreds of thousands of years ago.
- Brian Thomas and Jeffrey Tomkins, “How Reliable are Genomes from Ancient DNA?” Journal of Creation 28 no. 3 (2014): 92–98.
- Nathaniel T. Jeanson, “Mitochondrial DNA Clocks Imply Linear Speciation Rates Within ‘Kinds,’ ” Answers Research Journal 8 (2015): 273–304, https://answersingenesis.org/natural-selection/speciation/clocks-imply-linear-speciation-rates-within-kinds/.
- Nathaniel T. Jeanson, “Recent, Functionally Diverse Origin for Mitochondrial Genes from ~2700 Metazoan Species,” Answers Research Journal 6 (2013): 467–501, https://answersingenesis.org/genetics/mitochondrial-dna/recent-functionally-diverse-origin-for-mitochondrial-genes-from-~2700-metazoan-species/.
- Dennis Venema and Darrel Falk, “Does Genetics Point to a Single Primal Couple?” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/does-genetics-point-to-a-single-primal-couple.
- The 1000 Genomes Project Consortium, “A Global Reference for Human Genetic Variation,” Nature 526 (2015): 68-74, http://www.nature.com/nature/journal/v526/n7571/full/nature15393.html.
- Their offspring would have received 60 new mutations. So, 60 / 6,000,000,000 = 0.00000001%.
- Under the YEC model, there is no scientific reason to exclude mutations from happening after the entrance of sin into the world at the Fall. Instead, mutations likely played a minor role in generating the genetic diversity observable today — minor because of the sheer number of differences with which Adam and Eve were likely created.
- Robert W. Carter, “The Non-Mythical Adam and Eve! Refuting errors by Francis Collins and BioLogos,” http://creation.com/historical-adam-BioLogos; Nathaniel T. Jeanson and Jason Lisle, “On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity,” Answers Research Journal 9 (2016): 81–122, https://answersingenesis.org/natural-selection/speciation/on-the-origin-of-eukaryotic-species-genotypic-and-phenotypic-diversity/.
- Dennis Venema and Darrel Falk, “Does Genetics Point to a Single Primal Couple?” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/does-genetics-point-to-a-single-primal-couple.
- Specifically, the DNA sequences are called Alu sequences.
- The second claim addresses the arrangement and groupings of DNA differences along chromosome (technically termed linkage disequilibrium).
- Jianbin Wang et al., “Genome-wide Single-Cell Analysis of Recombination Activity and De Novo Mutation Rates in Human Sperm,” Cell 150 (2012): 402–412, http://www.cell.com/cell/abstract/S0092-8674%2812%2900789-1?_returnURL=http%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0092867412007891%3Fshowall%3Dtrue; Amy L. Williams et al., “Non-crossover Gene Conversions Show Strong GC Bias and Unexpected Clustering in Humans,” eLife 4 (2015): e04637, http://elifesciences.org/content/4/e04637; Pier Francesco Palamara et al., “Leveraging Distant Relatedness to Quantify Human Mutation and Gene-Conversion Rates,” Am. J. Hum. Genet. 97 (2015): 775–789.
- Jeanson and Lisle, “On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity,” p. 81–122.
- Ibid.
- Since Adam and Eve each would have been created with two versions of their 3 billion letter DNA sequence, and since Eve’s versions may have been slightly different than Adam’s, humanity may trace its genetic origins to 4 different starting points.
- Dennis Venema, “Mitochondrial Eve, Y-Chromosome Adam, and Reasons to Believe,” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/mitochondrial-eve-y-chromosome-adam-and-reasons-to-believe.
- See references in Nathaniel T. Jeanson, “A Young-Earth Creation Human Mitochondrial DNA ‘Clock’: Whole Mitochondrial Genome Mutation Rate Confirms D-loop Results,” Answers Research Journal 8 (2015): 375–378, https://answersingenesis.org/genetics/mitochondrial-genome-mutation-rate-/.
- Ibid.
- Nathaniel T. Jeanson, “Mitochondrial DNA Clocks Imply Linear Speciation Rates Within ‘Kinds,’ ” Answers Research Journal 8 (2015): 273–304, https://answersingenesis.org/natural-selection/speciation/clocks-imply-linear-speciation-rates-within-kinds/.
- About 1 mtDNA letter is mutated every ~6 generations, on average. See Nathaniel T. Jeanson, “A Young-Earth Creation Human Mitochondrial DNA ‘Clock’: Whole Mitochondrial Genome Mutation Rate Confirms D-loop Results,” Answers Research Journal 8 (2015): 375–378, https://answersingenesis.org/genetics/mitochondrial-genome-mutation-rate-/.
- Nathaniel T. Jeanson, “Recent, Functionally Diverse Origin for Mitochondrial Genes from ~2700 Metazoan Species,” Answers Research Journal 6 (2013): 467–501, https://answersingenesis.org/genetics/mitochondrial-dna/recent-functionally-diverse-origin-for-mitochondrial-genes-from-~2700-metazoan-species/; Nathaniel T. Jeanson, “Mitochondrial DNA Clocks Imply Linear Speciation Rates within ‘Kinds,’ ” Answers Research Journal 8 (2015): 273-304, https://answersingenesis.org/natural-selection/speciation/clocks-imply-linear-speciation-rates-within-kinds/; Nathaniel T. Jeanson, “A Young-Earth Creation Human Mitochondrial DNA ‘Clock’: Whole Mitochondrial Genome Mutation Rate Confirms D-loop Results,” Answers Research Journal 8 (2015): 375–378, https://answersingenesis.org/genetics/mitochondrial-genome-mutation-rate-/; Nathaniel T. Jeanson, “On the Origin of Human Mitochondrial DNA Differences, New Generation Time Data Suggest a Unified Young-Earth Creation Model and Challenge the Evolutionary Out-of-Africa Model,” Answers Research Journal 9 (2016): 123-130, https://answersingenesis.org/genetics/mitochondrial-dna/origin-human-mitochondrial-dna-differences-new-generation-time-data-both-suggest-unified-young-earth/.
- The range of numbers is due to the fact that the measured mutation rate has (like all biological data) a range of statistical uncertainty. Combined with the fact that there’s a range of generation times for humans (e.g., some women marry and bear children at age 15, others at age 35), we report a statistically reliable range of predictions for both creation and evolution.
- We used one of the shortest estimated time frames from the Flood to the present. Arguments could be made for longer time frames, but since our calculations with the shorter time frame already show agreement with current data, longer time frames would simply underscore the veracity of our results.
- Ibid.
- Nathaniel T. Jeanson, “Recent, Functionally Diverse Origin for Mitochondrial Genes from ~2700 Metazoan Species,” Answers Research Journal 6 (2013): 467–501, https://answersingenesis.org/genetics/mitochondrial-dna/recent-functionally-diverse-origin-for-mitochondrial-genes-from-~2700-metazoan-species/; Nathaniel T. Jeanson, “Mitochondrial DNA Clocks Imply Linear Speciation Rates within ‘Kinds,’ ” Answers Research Journal 8 (2015): 273–304, https://answersingenesis.org/natural-selection/speciation/clocks-imply-linear-speciation-rates-within-kinds/.
- Anon., “How Are the Ages of the Earth and Universe Calculated?”https://BioLogos.org/common-questions/scientific-evidence/ages-of-the-earth-and-universe/.
- For a recent discussion of the constant rate assumptions in astronomy, see: Jason Lisle, “Anisotropic Synchrony Convention — A Solution to the Distant Starlight Problem,” Answers Research Journal 3 (2010): 191–207, https://answersingenesis.org/astronomy/starlight/anisotropic-synchrony-convention-distant-starlight-problem/.
- The YEC community has even performed full-scale laboratory research projects to support this conclusion. For example, see Larry Vardiman, Andrew Snelling, and Eugene Chaffin, eds., Radioisotopes and the Age of the Earth, Vol. 2 (El Cajon, California: Institute for Creation Research; Chino Valley, Arizona: Creation Research Society, 2005).
- Dennis Venema, “Mitochondrial Eve, Y-Chromosome Adam, and Reasons to Believe,” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/mitochondrial-eve-y-chromosome-adam-and-reasons-to-believe.
- For example, see Max Ingman, et al., “Mitochondrial Genome Variation and the Origin of Modern Humans,” Nature 408 (2000): 708–713.
- E.g., they sequence the DNA from a parent-offspring pair (or perhaps even a grandparent-grandchild pair), and then count the number of differences that have arisen from ancestor to descendant. This represents one or more generational events; hence, units are reported in terms of mutations per generation.
- United Nations, Department of Economic and Social Affairs: Population Division, Fertility and Family Planning Section, “World Marriage Data 2012,” http://www.un.org/esa/population/publications/WMD2012/MainFrame.html; see also Nathaniel T. Jeanson, “On the Origin of Human Mitochondrial DNA Differences, New Generation Time Data Both Suggest a Unified Young-Earth Creation Model and Challenge the Evolutionary Outof- Africa Model,” Answers Research Journal 9 (2016): 123–130, https://answersingenesis.org/genetics/mitochondrial-dna/origin-human-mitochondrial-dna-differences-new-generation-time-data-both-suggest-unified-young-earth/.
- The 1000 Genomes Project Consortium, “A Global Reference for Human Genetic Variation,” Nature 526 (2015): 68–74, http://www.nature.com/nature/journal/v526/n7571/full/nature15393.html.
- Anjali G. Hinch et al., “The Landscape of Recombination in African Americans,” Nature 476 (2011): 170–175, http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3154982/.
- G. David Poznik et al., “Sequencing Y Chromosomes Resolves Discrepancy in Time to Common Ancestor of Males Versus Females,” Science 341 no. 6145 (2013): 562–565, http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4032117/.
- Dennis Venema, “Theory, Prediction and Converging Lines of Evidence, Part 3,” https://BioLogos.org/blogs/dennis-venema-letters-to-the-duchess/theory-prediction-and-converging-lines-of-evidence-part-3.
- Anon., “Creation Scientists and Other Specialists of Interest,” http://creation.com/creation-scientists.
- Anon., “Scientists and Belief,” http://www.pewforum.org/2009/11/05/scientists-and-belief/.
- Note that most of our references to evolutionary ideas come from the BioLogos website.
- For example, see chapter 23 of Douglas J. Futuyma, Evolution (Sunderland, MA: Sinauer Associates, Inc., 2013).
- Steven Benner, “Challenge or Preserve the Paradigm?” https://BioLogos.org/blogs/archive/challenge-or-preserve-the-paradigm.
- Darrel R. Falk, Coming to Peace with Science: Bridging the Worlds Between Faith and Biology (Downers Grove, IL: InterVarsity Press, 2004), p. 80–81.
- Michael Behe, “Correspondence w/ Science Journals: Response to Critics Concerning Peer-Review,” http://www.trueorigin.org/behe07.php.
- “The wicked in his proud countenance does not seek God; God is in none of his thoughts” (Ps. 10:4).
- Note that pride need not pervade every area of a person’s life. A Christian may be one of the most humble people you have ever met — in all areas but one, which happens to be the area in which he or she is currently undergoing sanctification.
- 92. All Christians, including the authors of this book, are susceptible to giving in to these two vices, as Scripture makes clear (e.g., Prov. 29:25 and John 12:42–43).
- Jeanson and Lisle, “On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity,” p. 81–122.



Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis