

Bacterial Life in Ancient Salt
Originally published in Journal of Creation 16, no 2 (August 2002): 110-113.
Living bacteria and dormant stages of bacteria known as spores, have been found in many sediments with reported ages of up to hundreds of million years. It is difficult to prove that these ancient bacteria were laid down during sedimentation (autochthonous origin) since it is also difficult to exclude the possibility that they entered the sediments later in time (allochthonous origin), or that they are sample contaminants.
Viable spores, with proven autochthonous origin, have recently been isolated from water inclusions in the late Permian Salado evaporite, New Mexico (USA). This evaporite has been dated at 250 Ma. However, current rates of bacterial life decay (the decrease in the number of viable spores with time) do not support the age of 250 Ma. Instead, these decay rates indicate an age for the Permian Salado salt layer of less than 35,000 years.
Age estimates based on bacterial decay rates are qualitative or semi-quantitative at best. Nevertheless, improved age estimates for the Salado evaporite might be possible by estimating the average temperature of the Salado evaporite and determining the decay rate of the bacterium Bacillus marismortui under conditions representative for water inclusions.
Present-day bacteria
Bacteria are present virtually everywhere on earth, many types surviving and even flourishing under extreme conditions such as pressurized water above 110°C, ice below -10°C, acids at pH values down to 2 and bases at pH values up to 13, and environments without oxygen, feeding on essentially everything including those inorganic nutrients available around mid-ocean hot springs. They even survive for long periods of time as spores without any nutrients.1
The presence of live bacteria in sediments has been the subject of study for almost a century. Autochthonous origin, the availability of nutrients over a long period of time, and contamination of the samples during sampling and handling are still subjects of dispute. During the ’70s, viable spores of a thermophilic (heat loving) bacterium were discovered in lake sediments with radiocarbon ages of 7 Ka or more. The likelihood of the sediments being contaminated with this bacterium later in time was excluded, as the sediments had never exceeded 35°C, the minimum temperature required for germination of this specific type of thermophilic bacterium. Another well-known example is the isolation of bacteria from the gut of a frozen Siberian mastodon, radio-dated at 11 Ka. These and other studies point to a maximum life-span for autochthonous bacteria in recent sediments of around 10,000 radiocarbon years, assuming the absence of significant metabolic activity.1
Ancient bacteria
The presence of bacteria in remote oil wells has been established, whereas the origin of these bacteria remains unsolved.
A point of controversy is the age of bacteria trapped in pre-Pleistocene Phanerozoic sediments. Once again the presence of bacteria is reported deep below the earth’s surface and in several types of deposits, including coal and oil layers and evaporites with reported ages up to 650 Ma. Evidence for autochthonous origin has not generally been accepted, as it is difficult to exclude allochthonous origin or contamination during sampling and handling.1,2,3 For example, the presence of bacteria in remote oil wells has been established, whereas the origin of these bacteria remains unsolved. These may have been trapped during the formation of the oil or have been transported by aquifer activity, or they may have migrated over tens to hundreds of millions of years.4
Recently, viable Bacillus sphaericus spores were isolated from an extinct bee species enclosed in amber with a reported age of 25–30 Ma.5 The careful selection of the sample and thorough precautions taken against contamination has made this a convincing find.2
Even more recently, ancient bacteria have been isolated from autochthonous water inclusions in salt samples from the Permian Salado evaporite (New Mexico) with a reported age of 250 Ma.2,6 Based on 16S ribosomal DNA comparison, these ancient bacteria show remarkable similarity to Bacillus marismortui, (named after the Dead Sea) and Virgobacillus pantothenticus. The difference with V. pantothenticus is small, with an homology of 97.1% (although Vreeland et al indicate 97.5% similarity). Of the 1560 base pairs, 45 are different, which corresponds to 1x10-10 substitutions per base pair each year. This substitution rate is small but possibly acceptable.7 However, the 99% homology with B. marismortui is even more remarkable. If these ancient bacteria are indeed relatives of B. marismortui, which is yet to be established, how can this difference be so small? The free living B. marismortui have supposedly experienced 250 million years of mutations and a much larger difference to their isolated counterparts would be expected,2,6 although an efficient DNA repair mechanism during germination could partly explain the similarity.8
In this respect, the ultimate life-span of isolated bacteria is a key parameter which will be discussed in more detail below.9
Bacterial survival
In theory, bacteria can live indefinitely as long as a sufficient supply of nutrients are available. They multiply asexually, whereby one cell simply splits into two cells, and both mother and daughter cells (which in fact are indistinguishable) are rejuvenated and the population remains constantly viable so long as optimal nutrition is available and the levels of excreted toxic substances is low. The life of bacteria is limited in the absence of nutrients, but it is impossible to determine their life expectancy. Nevertheless, the number of viable individuals steadily decreases over time as the level of nutrients decrease and the levels of toxic excretions increase. The decay rate of spores is usually expressed as the decimal reduction time, D(10), which is the time required, at a given temperature, for the number of viable bacteria in a population to be reduced to 10%. This rate is comparable with that from other exponential decay processes such as first-order chemical reactions or radioactive decay.
Bacteria can typically be stored for about one year if dried under vacuum, and for several years at liquid nitrogen temperature (-196°C).10 They can survive for much longer if they can maintain basal metabolic activity. Bacterial cultures have been found in Siberian permafrost at an average temperature of around -10°C. The permafrost is estimated at between one thousand and one million years. The suggestion has been made that despite the low temperature, the bacteria must have been active, otherwise they could not have survived for so long.11 Supporting evidence for low-temperature activity is the recent discovery of bacteria in snow and ice in the surface layers of Antarctica, which indeed have metabolic activity at -12°C to -17°C.12
It is interesting to note that the Antarctica researchers discovered members of Deinococcus, a genus of bacteria that was already known in the preserved food industry for its ability to withstand ionizing radiation, including X-rays and radioactivity.13 This is remarkable, as such high irradiations are not present in the natural environment, and it was unclear what advantage this ability could give. The answer could be that in the Antarctica snowfields, Deinococcus species are subject to intensive UV irradiation which causes ionisation.12 This example again illustrates the remarkable ability of some bacteria to survive under extreme conditions.
Most bacteria find it difficult to survive when subjected to high temperature, radiation (including UV irradiation), acidic or basic environments, oxidizing agents (including oxygen), certain inorganic compounds, mechanical stress and wet conditions.10 Nevertheless, many of these conditions can be favourable for bacteria if they occur in a water inclusion in an evaporite, even if water is present. Although salt is detrimental to most bacteria, some such as B. marismortui can survive as spores or even flourish under saline conditions below 25% w/v.
In summary, some bacteria can withstand severe conditions as long as metabolic activity can take place. In the absence of nutrients, the life-time of active bacteria could be years or decades, but survival over thousands of years in the absence of metabolic activity under otherwise optimal conditions is already challenged. Can bacteria imprisoned in a water inclusion in salt survive over 250 Ma?
The survival of spores
Some bacteria, including Bacillus species, can form spores in the absence of nutrients or when under stress. Spores are inactive dormant stages which tide bacteria over a harsh period of time, and include cysts, exospores and myxospores which are less stable than the Bacillus endospores10 considered here. The transition of a bacterium to a spore is a complex process resulting in a nucleus of stable compacted DNA and enzymes, wrapped in an impermeable keratin-like skin. There is hardly any water, but a large amount of the conservation complex calcium-dipicolinate is present to keep the enzymes inactivate and to stabilize the nucleic acids. The DNA is also protected by a group of unique DNA-binding proteins, called a/b-type SASP.13 Most bacteria cannot survive heating at 80°C for 10 minutes (pasteurization), but a few spores do, some even lasting for hours in boiling water.

Table 1. Life decay constants of several bacterial spores expressed as times of decimation, D(10), in seconds.
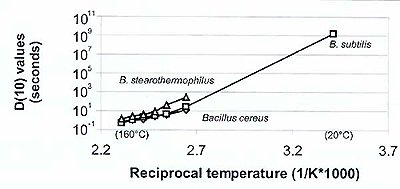
Figure 1. Arrhenius plot of bacterial life decay.
How long can spores survive? Research conducted on the soil residues on plant roots stored under dry conditions in a herbarium gives us some indication. After 200–300 years, some spores from B. subtilis and B. lichenifornis were still viable. In addition, B. coagulans and B. circulans were identified in 50 to 100 year-old samples. This was equivalent to a D(10) of 50 years for B. subtilis and B. lichenifornis, and means that after 1,000 years only a few viable spores from the original large number of bacteria can still be expected from 1,000 kg of soil.10
D(10) times have been reported for various temperatures, including sterilization conditions (Table 1). The sterilization stability of B. stearothermophilus is exceptional and probably not representative for most other Bacillus species. Unfortunately, data for B. marismortui under conditions characteristic for water inclusions in salt is not available. B. marismortui does not survive laboratory sterilization precautions,6 but this feature cannot be translated into a D(10) time. Figure 1 is a plot of the D(10) data from Table 1 on a logarithmic scale as a function of the reciprocal temperature (Arrhenius plot), and provides the trend of D(10) values as a function of temperature.
The ‘bacterial age’ of the Salado evaporite
Bacillus marismortui forms spores when the salt concentration exceeds 25% w/v. Under saturated saline conditions, spores are not expected to ‘wake up’ regardless of the nutrients available. The number of viable spores in the Salado deposit has declined steadily with time, from possibly 100 million bacteria per ml originally2 to only a few viable spores in just three out of 66 inclusions, with a total volume of 0.365 ml (for the 66 inclusions) having been evaluated to date.6 Assuming a minimum of three viable spores in 0.365 ml liquid means that seven periods of decimation have passed. Taking a D(10) of 50 years results in a ‘bacterial age’ of 350 years for the Salado evaporite. Additional supporting evidence for the young age of this evaporite is the previously mentioned similarity between the 16S ribosomal DNA of the ancient bacteria and B. marismortui or V. pantothenticus.
The difference with the 250 Ma reported for the Salado evaporite is almost six orders of magnitude! An unreasonable D(10) of 35 Ma instead of just 50 years is required to reach a total of 250 Ma. Conversely, assuming the evaporite has an age of 250 Ma and a decimation time of 50 years implies that the original bacteria have experienced 5 million decimation periods. It is impossible to find a living organism after 5 million decimations.
An age of 350 years for the Salado deposit is within historic time and of course not realistic. There are a number of uncertainties, that must be considered.
The small number of viable spores (three as a minimum) has hardly any statistical significance and results in a wide range of possible ages, with greater ages more probable than shorter ones.
The D(10) of B. marismortui under conditions representative of water inclusions is unknown and needs to be established independently.
The times of decimation for different species might vary by one order of magnitude (Table 1).
A temperature change of 25°C has an effect of two orders of magnitude on the time of decimation (Fig. 1).
An average evaporite temperature at or below 0°C over time is highly improbable.
An average temperature above 20°C might be more realistic and results in shorter D(10) times and lower ages.
The calculated ‘bacterial age’ of 350 years is therefore highly uncertain and can easily vary by two, or possibly three, orders of magnitude with greater ages more probable than younger ages. This results in an estimated range of 200 to 35,000 years, corresponding to 4 log cycles of 50 years at 20°C or 7 log cycles and a D(10) of 5000 years at 0°C, respectively. It is difficult to imagine conditions leading to 350,000 years, but even this estimate differs by more than two orders of magnitude with the 250 Ma reported for the Salado evaporite. This gap may even increase when claims of 650 million-year-old bacteria in evaporites are confirmed.
Surprisingly, the presence of viable autochthonous bacterial spores in the Salado evaporite has not led to a challenge of its geological age. Parkes assumes virtual bacterial immortality rather than challenging the geological time scale. Even survival of bacteria in meteorites and the origin of life on earth via inter-planetary transport of bacterial spores (‘panspermia’) are being considered.2 More recently, Lindahl has denied the high age of these bacteria based on the impossibility of DNA surviving longer than 100,000 years, stating that the recovered bacteria are common extant ones.14
Conclusions
The presence of viable spores of bacteria in water inclusions in the Permian Salado evaporite, in combination with bacterial decay rates, is incompatible with the age of 250 Ma reported for this evaporite.
The ‘bacterial age’ of the Permian Salado evaporite has here been estimated semi-quantitatively and is probably within the range of 350 and 35,000 years, under the assumption that decay rates available from literature are representative for inclusion conditions.
Estimates of the average temperature of the evaporite over time and measurement of times of decimation for Bacillus marismortui within a wide range of temperatures are key parameters for improving the ‘bacterial age method’ estimate.
Footnotes
- Postgate, J., The outer reaches of life, University Press, Cambridge, 1994.
- Parkes, R.J., A case of bacterial immortality? Nature, 407:844–845, 2000.
- Roth, A.A., Origins, Review and Herald Publishing Association, Hagerstown, p. 164, 1998.
- Magot, M., Similar bacteria in remote oil fields, Nature, 379:681, 1996.
- Cano, R.J. and Borucki, M., Revival and identification of bacterial spores in 25–40 million year old Dominican amber, Science, 268:1060–1064, 1995.
- Vreeland, R.H., Rosenzweig, W.D. and Powers, D.W., Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal, Nature, 407:897–900, 2000.
- Hartl, D.L. and Clark, A.G., Principles of Population Genetics, Sinauer, Sunderland, p. 107, 1989.
- Nicholson, W.L., Munakata, N., Horneck, G., Melosh, H.J. and Setlow, P., Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments, Microbiol. and Molec. Biol. Rev., 64:548–572, 2000.
- Van der Heide, E., Leven in aardlagen, Bijbel en Wetenschap, 225:5–8, 2000.
- Schlegel, H.G., General Microbiology, 7th Edition, Cambridge, 1993.
- Shi, T., Reever, R.H., Gilchinsky, D.A. and Friedman, E.I., Characterisation of viable bacteria from Siberian Permafrost by 16S rDNA sequencing, Microb. Ecol., 33:169–179, 1997.
- Carpenter, E.J., Lin, S. and Capone, D.G., Bacterial Activity in South Pole Snow, Appl. Env. Microb., 66:4514–4517, 2000.
- Makarova, K.S., Aravind, L., Wolf, Y.I., Tatusov, R.L., Minton, K.W., Koonin, E.V. and Daly, M.J., Genome of the extremely radiation-resistant bacterium Deinococcus radiodurans viewed from the perspective of comparative genomics, Microbiol. Molec. Biol. Rev., 65:44–79, 2001.
- Lindahl, T., Unlocking nature’s ancient secrets; review of: The Molecule Hunt: Archaeology and the Search for Ancient DNA by Martin Jones, Nature 413(6854):358–359, 2001.
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis