
Chapter 24
Vestigial Organs—Evidence for Evolution?
by
Dr. David Menton
on
July 7, 2014
Republished with permission and featured in
The New Answers Book 3
Vestigial organs have long been one of the classic arguments used as evidence for evolution. The argument goes like this: living organisms, including man, contain organs that were once functional in our evolutionary past, but that are now useless or have reduced function. This is considered by many to be compelling evidence for evolution. More importantly, vestigial organs are considered by some evolutionists to be evidence against creation because they reason a perfect Creator would not make useless organs.
The word vestige is derived from the Latin word vestigium, which literally means a “footprint.” The Merriam-Webster’s Dictionary defines a biological vestige as a “a bodily part or organ that is small and degenerate or imperfectly developed in comparison to one more fully developed in an earlier stage of the individual, in a past generation, or in closely related forms.”
Darwin on “Rudimentary Organs”
Darwin speculated that these rudimentary organs once served a function necessary for survival, but over time that function became either diminished or nonexistent.
Charles Darwin was perhaps the first to claim vestigial organs as evidence for evolution. In chapter 13 of his Origin of Species, Darwin discussed what he called “rudimentary, atrophied and aborted organs.” He described these organs as “bearing the plain stamp of inutility [uselessness]” and said that they are “extremely common or even general throughout nature.” Darwin speculated that these rudimentary organs once served a function necessary for survival, but over time that function became either diminished or nonexistent.
In Darwin’s book The Descent of Man, he claimed about a dozen of man’s anatomical features to be useless including the muscles of the ear, wisdom teeth, the appendix, the coccyx (tailbone), body hair, and the semilunar fold in the corner of the eye. To Darwin, this was strong evidence that man had evolved from primitive ancestors.
The List of “Vestigial Organs” Grows
In 1893 the German anatomist Robert Wiedersheim expanded Darwin’s list of “useless organs” to 86. Listed among Wiedersheim’s “vestigial” organs were such organs as the parathyroid, pineal and pituitary glands, as well as the thymus, tonsils, adenoids, appendix, third molars, and valves in veins.1 All of these organs have been subsequently shown to have useful functions and indeed some have functions essential for life.
Wiedersheim’s vestigial organs were presented as one of the so-called “proofs” of evolution in the famous Scopes “Monkey Trial” of 1925. Horatio Hackett Newman, a zoologist from the University of Chicago, stated on the witness stand that “there are, according to Robert Wiedersheim, no less than 180 [sic] vestigial structures in the human body, sufficient to make a man a veritable walking museum of antiquities.”2
Vestigial Organs Still Used as Evidence for Evolution
For over 100 years, evolutionists have continued to use vestigial organs as evidence for evolution. In 1971 the Encyclopedia Britannica claimed there were more than 100 vestigial organs in man, and even as recently as 1981, some biology textbook authors were claiming as many as 100 vestigial organs in the human body.3 One of the most popular current biology textbooks declares that “many species of animals have vestigial organs.” Examples cited in humans include the appendix, “tailbone,” and muscles that move the ear.4
In addition to textbooks, countless popular science magazines, evolution blogs, and websites continue to promote vestigial organs as evidence for evolution. A website sponsored by the Discovery Channel, for example, assures us that “the human body has something akin to its own junk drawer,” and that this junk drawer “is full of vestigial organs, or souvenirs of our evolutionary past.”5
Problems with Vestigial Organs as Evidence for Evolution
Why Do Useless Organs Persist?
Darwin himself pointed out a flaw in the vestigial organ argument. He wondered how once an organ is rendered useless, it can continue to be further reduced in size until the merest vestige is left. In chapter 14 of Origin of Species he declared, “It is scarcely possible that disuse can go on producing any further effect after the organ has once been rendered functionless. Some additional explanation is here requisite which I cannot give.” Why, indeed, would useless organs continue to exist for millions of years after they ceased to have any selective advantage?
The Loss of Useful Organs Doesn’t Explain Their Origin
A problem for using vestigial organs as evidence for “amoeba to man” evolution is that the chief burden of the macro evolutionary explanation is to account for the spontaneous origin of new functional organs—not the loss of functional organs. While evolution might require the loss of functional organs, it is the acquisition of fundamentally new organs that remains unexplained by random mutations and natural selection.
How Can We Be Certain an Organ Is Useless?
The problem with declaring any organ to be without function is discriminating between truly functionless organs and those that have functions that are simply unknown. Indeed, over the years nearly all of the organs once thought to be useless have been found to be functional. When we have no evidence for the function of an organ, it is well to bear in mind that absence of evidence is not evidence of absence.
Declaring Useful Organs to Be Useless Can Be Dangerous
Once an organ is considered to be useless, it may be ignored by most scientists, or even worse, surgically removed by physicians as a useless evolutionary leftover. The oft repeated claim that the human appendix is useless is a case in point. The evolutionist Alfred Romer in his book The Vertebrate Body said of the human appendix: “Its major importance would appear to be financial support of the surgical profession.”6 We can only wonder how many normal appendices have been removed by surgeons since Darwin first claimed them to be a useless vestige. Even more frightening would be the surgical removal of a “useless” parathyroid or pituitary gland.
The Definition of Vestigial Organs Has Been Changed
As the list of “functionless” organs has grown smaller and smaller with advancing knowledge, the definition of vestigial organs has been modified to include those whose functions are claimed to have “changed” to serve different functions. But such a definition removes the burden of proof that vestigial organs are a vestige of evolution. Thus, the evolutionist might concede that the human coccyx (“tail bone”) does indeed serve an important function in anchoring the pelvic diaphragm—but still insist, without evidence, that it was once used by our ancestors as a tail.
Circular Reasoning
The most conspicuous logical flaw in the use of vestigial organs as evidence for evolution is circular reasoning.
The most conspicuous logical flaw in the use of vestigial organs as evidence for evolution is circular reasoning. Evolutionists first declare vestigial organs to be a result of evolution, and then they turn around and argue that their existence is evidence for evolution. This kind of argument would hardly stand up in a court of law.
There Are Other Explanations for Vestigial Organs
Vestiges of Embryology
Evolutionists insist on explaining vestigial organs only in terms of evolution, but other explanations are more plausible and even provable. For example, the human body does have many organs and structures that are clearly vestiges of our embryological development. While it is quite easy to prove that an organ or structure is a vestige of embryology, there can be no empirical evidence to support the speculation that an organ is a vestige of evolution.
There are several structures that function during the development of the embryo and fetus that appear to be no longer used after birth. Remnants of these once-functional structures persist throughout life. Such structures perfectly fit the definition of a vestige, but they are not vestiges of evolution. The following are a few examples of embryological vestiges.
Ligamentum arteriosum—obliterated remnant of the ductus arteriosus, an artery that shunted blood from the pulmonary trunk to the descending aorta, thus bypassing the lung during fetal development. In certain cases of congenital heart defects, the ductus arteriosus actually continues to function for some time after birth to keep the baby alive.
Ligamentum teres hepatis—obliterated remnant of the umbilical vein that shunted much of the oxygenated blood away from the liver to the inferior vena cava during fetal development.
Median umbilical ligament—an obliterated vestige of the allantois, a pouch extending off of the embryonic cloaca. The allantois disappears very early in gestation after functioning as a scaffolding to help construct the umbilical cord; this remnant is seen as a ligament extending from the bladder to the umbilicus (bellybutton).
Sexual Dimorphism
In most primates there are striking anatomical differences between males and females of the same species. These differences between the sexes are refered to as sexual dimorphism. The skulls of a male and female gorilla, for example, might not be recognized as from the same species if one had never seen them in the flesh. The difference between the sexes is not as dramatic in the case of humans, though they are dimorphic. The bodies of human males and females differ mostly in the organs related to reproduction.
Up until the end of the sixth week of embryological development, the reproductive organs of males and females are indistinguishable. After this time, the genital organs of both sexes develop from the same common starting tissues under the control of sex chromosomes (XX in the female and XY in the male) and various hormones. As a result of their embryological development from the same primordia, each sex contains vestigial components of the other sex.
Almost every organ of the female reproductive system can be found in a different or vestigial form in the male reproductive system (and vice versa). For example, in the male, the prostatic utricle (an out pouching of the prostatic urethra having no known function) is a remnant of the paramesonephric duct that develops into the uterus and oviducts of the female. Clearly, the vestigial organs of reproduction are not a result of evolution but rather embryological development.
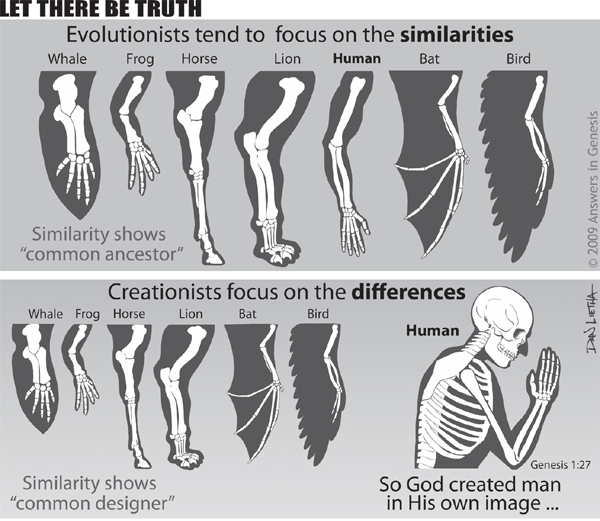
Homology
Many vestigial organs are examples of homology but not necessarily of evolution.
Many vestigial organs are examples of homology but not necessarily of evolution. Homology is an underlying similarity between different kinds of animals recognized by both evolutionists and creationists. All terrestrial vertebrates, for example, share a widespread similarity (homology) of body parts. Evolutionists insist that this similarity is the result of evolution from a common ancestor. Creationists, on the other hand, argue that this similarity reflects the theme of a common Creator and the need to meet similar biological requirements.7
For example, all vertebrates with true limbs (amphibians, reptiles, birds, and mammals) have the same basic limb structure at least during their embryological development. This standard vertebrate limb consists of an upper limb comprising one bone, a lower limb comprising two bones, and a hand or foot bearing five digits (fingers and toes). Thus, the limbs of all limbed vertebrates share fundamental similarities, with each being specialized to meet the needs of each species.
Horses have five digits while developing as an embryo, but generally all but one (the third digit) is absorbed before birth. Vestiges of the second and third metacarpal (and metatarsal) bones are visible in the modern horse as the splint bones. Some fossil horses, however, had three toes, but both three-toed and one-toed horses have been found together in the fossil record. In National Geographic magazine, for example, there is a picture of the feet of both a three-toed horse (Pliohippus) and a one-toed horse (Equus) that were found at the same volcanic site in Nebraska.8
Human hair is an example of a homologous structure declared to be vestigial by evolutionists. All mammals have hair. Hair may vary from the compacted hairs of a rhinoceros horn to the quills of a porcupine. To declare the unique hairs of one mammal to be vestigial to those of another is biological nonsense.
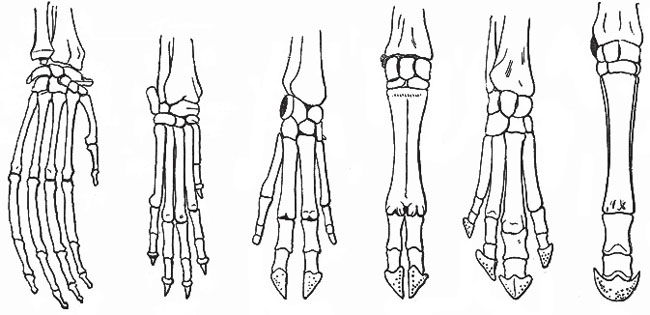
Homology in vertebrate limbs does not prove they came from a common ancestor.
Evaluating Currently Claimed “Vestigial” Organs
It may prove useful for the reader to use the forgoing discussion of vestigial organs to evaluate some current claims for such structures. The website LiveScience lists what it regards as the top ten “vestigial” organs.9 Five of these are found in humans, and are discussed below in order of their perceived importance by LiveScience.
The Appendix
Ever since Darwin, the appendix has been the prime example of a “useless” organ. LiveScience says of the appendix that “it is a vestigial organ left behind from a plant-eating ancestor.” In the middle of the 20th century, surgeons often removed the appendix electively during abdominal surgery, assuming it had no function. According to most evolutionists, the appendix is a vestige of the caecum (an expanded area at the beginning of the large intestine) left over from our plant-eating ancestors. But since humans have a well-developed caecum as well as an appendix, the appendix can hardly be considered a vestigial caecum. In his book The Vertebrate Body, evolutionist Alfred Romer said that the appendix is “frequently cited as a vestigial organ supposedly proving something about evolution. This is not the case. . . .”10
The important point is that the presence or absence of an appendix (or a caecum) reveals no evolutionary pattern whatever. An appendix is not found in any invertebrate, amphibian, reptile, or bird. Only a few diverse mammals have an appendix.
The appendix is found, for example, in rabbits and some marsupials such as the wombat, but is not found in dogs, cats, horses, or ruminants. Both Old World and New World monkeys lack an appendix, while anthropoid apes and man have an appendix.11 The appendix is a complex, highly specialized organ with a rich blood supply—not what one would expect from a vestigial organ. The appendix is part of the gut associated lymphoid tissue (GALT), and has long been suspected of playing an immunological role much like that of the tonsils and adenoids (also once considered to be vestigial).
Recent evidence suggests that the appendix is well suited to serve as a “safe house” for commensal (mutually beneficial) bacteria in the large intestine. Specifically, the appendix is believed to provide support for beneficial bacterial growth by facilitating re-inoculation of the colon with essential bacteria in the event that the contents of the intestinal tract are purged following exposure to a pathogen.12
Male Breast Tissue and Nipples
It is surprising that evolutionists still continue to bring up the matter of the male breast (mammary gland) as a vestigial organ. Are they proposing that the males once nursed the young early in their evolution but no longer do so? Of course not. So how then does the evolutionist explain the male’s vestigial mammary gland if it is not a consequence of evolution?
Vestigial mammary glands in males can only be understood in terms of embryology—not evolution. Mammary glands begin to develop in both males and females in the sixth week of gestation. At the time of birth, the rudimentary mammary glands of males and females are identical. In fact, both male and female mammary glands may be slightly enlarged at birth and secrete a fluid that is commonly known as “witches milk.” This results from hormones that induce milk production in the mother being passed through the placenta to the fetal circulation.13
The male mammary gland is clearly a rudimentary or vestigial structure, but even the mammary gland of the nonlactating female might be considered vestigial. Female mammary glands are never fully developed and functional except during times of breast feeding the young. Should the evolutionist then consider the nonlactating female mammary gland to also be a vestige of evolution? The old evolutionist axiom that “nothing in biology makes sense except in the light of evolution” might better say that nothing in biology makes sense in the light of evolution.
Wisdom Teeth
Darwin was the first to popularize the notion that wisdom teeth are vestigial leftovers from our ape-like ancestors. The inherent racism of Darwinism is apparent when in his Descent of Man, Darwin declared that wisdom teeth are often lacking in “the more civilized races of man” in contrast to the “melanin (black) races where the wisdom teeth are furnished with three separate fangs, and are generally sound.”14
Wisdom teeth, properly known as third molars, generally appear between the ages of 15 and 27 in both the upper and lower jaws of man. Many evolutionists consider them to be vestigial because unlike apes, third molars often fail to develop properly in man due to lack of space in the jaw. They argue that apes with their sloping face have longer jaws than man, and that when ape-like creatures evolved into humans with a vertical face and shorter jaws, there was no longer room for third molars.
Third molars are hardly useless vestiges. When there is adequate room for their development, they are fully functional molars and are used in chewing much as the first and second molars. Thinking them to be vestigial, many dentists in the past routinely removed third molars whether or not they were causing problems. It has been estimated that in America, only 20 percent of all young people with otherwise healthy teeth develop impacted third molars that require medical attention, while in the past, nearly nine out of ten American teenagers with dental insurance had their third molars extracted.15
The “Tailbone” (Coccyx)
The so-called “tailbone” is perhaps the most commonly touted example in man of a “useless” evolutionary vestige. According to evolutionary dogma, the tailbone, properly called the coccyx (because of its similarity to the shape of a cuckoo’s beak), is a vestigial tail left over from our tailed monkey-like ancestors. Once again, many in the medical profession have been taken in by evolutionary speculation but mercifully, they have refrained from surgically removing the normal coccyx.
Even human abnormalities that have nothing to do with the coccyx have been declared to be “human tails.” In a report in The New England Journal of Medicine, titled “Evolution and the Human Tail,” Ledley described a two-inch long fleshy growth on the back of a baby, which he claimed to be a “human tail,” though he conceded that it showed none of the distinctive biological characteristics of a tail! In fact, the “tail” was merely a fatty outgrowth of skin that wasn’t even located in the right place on the back to be a tail! Still, Ledley declared that “even those of us who are familiar with the literature that defined our place in nature (Darwinism)—are rarely confronted with the relation between human beings and their primitive ancestors on a daily basis. The caudal appendage brings this reality to the fore and makes it tangible and inescapable.”16
The human coccyx is a group of four or five small vertebrae fused into one bone at the lower end of our vertebral column. The coccyx is commonly called the “tailbone” because of its superficial similarity to a tail. The coccyx does occupy the same relative position at the end of our vertebral column as does the tail in tailed primates, but then, where else would it be? The vertebral column is a linear row of bones that supports the head at one end and the other must end somewhere. Wherever it ends, evolutionists will be sure to call it a vestigial tail.
Many modern biology textbooks give the erroneous impression that the human coccyx has no real function other than to remind us of our evolutionary ancestry. In fact, the coccyx has some very important functions. Six muscles converge from the ring-like bones of the pelvic brim to anchor on the coccyx, forming a bowl-shaped muscular floor of the pelvis called the pelvic diaphragm. The incurved coccyx with its attached pelvic diaphragm supports the organs in our abdominal and pelvic cavities such as the urinary bladder, uterus, prostate, rectum, and anus. Without this critical muscular support, these organs could be easily herniated. The urethra, vagina, and anal canal pass through the muscular pelvic diaphragm, and thus the diaphragm serves as a sphincter for these structures.
Erector Pili and Body Hair
Evolutionists have long insisted that human body hair, and the small muscles (erector pili) attached to these hairs, are useless vestiges from our hairy ancestors. But human hair is as fully functional as that of any other mammal.
The body of man, like that of most mammals, is covered with hairs except for the palms and soles. But man, unlike other mammals, has mostly tiny colorless hairs called vellus hairs covering the seemingly “unhaired” parts of his body. This gives humans the appearance of being “hairless” with the exception of such areas as the scalp, axilla, chest, and genital regions. But in fact, if we count the tiny vellus hairs, humans have about as many hairs per square inch on their nose and forehead as they do on the top of their head. Indeed, hair density per square inch is approximately the same on the human body as it is for most primates.
Hair grows from tube-like structures in the skin called hair follicles. Most hair follicles are capable of making more than one type of hair depending in part on age, location, and hormonal stimulation. The first hairs to grow from the follicles of the developing baby are long silky hairs called lanugo hairs. These hairs, which cover most of the body, are usually shed before birth and are replaced with tiny vellus hairs. Thus, the newborn baby may appear to be mostly hairless, but in fact is covered with vellus hairs.
The long pigmented hairs on our scalp and elsewhere on our body are called terminal hairs. Terminal hairs grow from follicles that once produced lanugo and vellus hairs and with age may be replaced once again with vellus hairs. For example, after a boy reaches sexual maturity he may begin to lose terminal scalp hairs, which are replaced with vellus hairs, giving the appearance of baldness. Conversely, some vellus hairs on the face may be replaced with terminal hairs, producing a beard.
Evolutionists argue that human body hairs are vestigial (useless) because there are so few long terminal hairs compared to tiny vellus hairs. Hair serves as thermal insulation in most mammals, which is important because most animals are incapable of regulating their body temperature by sweating. Man, on the other hand, is a profuse sweater and can maintain body temperature over a much wider range of ambient temperature than nearly all other mammals. Long body hair of the type seen on most mammals would interfere with the evaporative water loss necessary for human thermoregulation by sweating.
In most mammals, hair serves as an important barrier to ultraviolet radiation from the sun. While human scalp hair serves a similar function on the typically exposed top of our head, our primary defense against UV damage is tanning and wearing clothes.

@ Tyler Olson, Dreamstime.com
Goose bumps are not remnants of an evolutionary past, but serve several functions for humans.
An important function of hair is its sensory function. All hair follicles, regardless of size, are supplied with sensory nerves so that they may be considered to be mechanoreceptors. Our hairs are like small levers that, when moved by any physical stimulus including air, send sensory signals to our brain. This is true of both the tiny vellus hairs and the long terminal hairs. This sensory function of hair can hardly be considered vestigial. Another important function of hair follicles is the restoring of the epidermal skin surface following cuts and deep abrasions. Human hair follicles, regardless of size, serve as an important source of epidermal cells for recovering the skin’s surface (reepithelialization) when broad areas of the epidermis are lost. If it were not for man’s abundant hair follicles and sweat ducts, even routine skin abrasions might require a skin transplant.
All hairs are associated with muscles, and most have a muscle called the erector pili, which serves to move the hair from its normal inclined position to a more erect position. In the case of the vellus hairs of man this produces what is commonly called “goose bumps.” This muscle is in a position to help squeeze oil from the sebaceous glands, which are also attached to the hair follicle. Erector pili muscles are supplied with nerves of the sympathetic nervous system, which is often associated with our response to “flight and fright” stimuli. Thus, when we are frightened we may get goose bumps. We also get goose bumps when we are chilled. Contraction of the erector pili muscles produces heat, and if this response is inadequate to warm the body, shivering may follow, which involves repeated contractions of the large body muscles.
Is the Argument for Vestigial Organs Vestigial?
Over the years, advancement in our understanding of biological science has raised serious doubts about vestigial organs as evidence for evolution. Creationists have subjected the evolutionary interpretation of vestigial organs to strong criticism.17 Even some evolutionists are now urging that vestigial organs be downplayed or even abandoned as evidence for evolution. The evolutionist S.R. Scadding, for example, has critically examined vestigial organs as evidence for evolution. He concluded: “Since it is not possible to unambiguously identify useless structures, and since the structure of the argument used is not scientifically valid, I conclude that ‘vestigial organs’ provide no special evidence for the theory of evolution.”18 But like the long discredited recapitulation myth (that embryos pass through stages of their evolutionary history), vestigial organs continue to be used as evidence for evolution.
The New Answers Book 3

Do you have answers to the big questions about the Christian faith, evolution, creation, and the biblical worldview?
Read Online Buy BookFootnotes
- R. Wiedersheim, The Structure of Man: An Index to His Past History (London: Macmillan and Co., 1895).
- The World’s Most Famous Court Trial (Dayton, TN: Bryan College, 1990). This book is a word-for-word transcript of the famous court test of the Tennessee Anti-Evolution Act, at Dayton, July 10 to 21, 1925, including speeches and arguments of attorneys, testimony of noted scientists, and Bryan’s last speech.
- S.R. Scadding, “Do Vestigial Organs Provide Evidence for Evolution?” Evolutionary Theory 5 (1981): 173–176.
- K.R. Miller and J. Levine, Biology: Teachers Edition (Upper Saddle River, NJ: Pearson Prentice Hall, 2006), p. 384.
- www.health.howstuffworks.com/vestigial-organ.htm/printable.
- A. S. Romer and T. S. Parsons, The Vertebrate Body (Philadelphia: Saunders College Publishers, 1986), p. 389.
- G.E. Parker, Creation: Facts of Life (Green Forest, AR: Master Books, 2006), p. 43–53.
- M.R. Voorhies, “Ancient Ashfall Creates a Pompeii of Prehistoric Animals,” National Geographic, January 1981, p. 74.
- www.livescience.com/animals/top10_vestigial_organs.html.
- Romer and Parsons, The Vertebrate Body, p. 358.
- J.W. Glover, “The Human Vermiform Appendix: A General Surgeon’s Reflections,” Technical Journal 3 no. 1 (1988): 31–38.
- R.R. Bollinger et al., “Biofilms in the Large Bowel Suggest an Apparent Function of the Human Vermiform Appendix,” Journal of Theoretical Biology 249 no. 4 (2007): 826–831.
- K.L. Moore, The Developing Human (Philadelphia, PA: W.B. Saunders Company, 1988), p. 427.
- C. Darwin, The Descent of Man and Selection in Relation to Sex (New York, NY: D. Appleton and Company, 1896), p. 20.
- A.J. MacGregor, The Impacted Lower Wisdom Tooth (New York, NY: Oxford University Press, 1985); a good review of wisdom teeth and the consequences of considering them to be evolutionary vestiges may be found at https://answersingenesis.org/human-body/vestigial-organs/are-wisdom-teeth-evidence-evolution/
- F.D. Ledley, “Evolution and the Human Tail: A Case Report.” N Engl J Med 306 no. 20 (1982): 1212–1215.
- J. Bergman and G. Howe, Vestigial Organs are Fully Functional (Terre Haute, IN: Creation Research Society Books, 1990).
- S.R. Scadding, “Do Vestigial Organs Provide Evidence for Evolution?” Evolutionary Theory 5 (1981): 173.
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis