

Irreducible Complexity: Some Candid Admissions by Evolutionists
Originally published in Journal of Creation 17, no 2 (August 2003): 56-59.
Although some evolutionists try to deny the existence of irreducible complexity, others, while using different wording, tacitly admit that it is a serious problem for organic evolution. Three intertwined examples of irreducible complexity discussed in this brief report are 1) The origin of novel regulatory complexes governing gene behavior, 2) The hoped-for evolution of genes that have novel functions relative to their supposedly ancestral genes, and 3) The origin of new proteins that have a very different function from the presumably ancestral proteins. In each case, evolutionists point to instances of simultaneous changes in gene expression. However, the observed phenotypic effects are always small. The simultaneous appearance of several mutations, even if neutral or beneficial, is not yet proof that any combination of them can produce even one new irreducibly complex system.
Living things are extremely complex. Evolutionary theory rests upon the premise that all biological systems could have evolved from progressively simpler systems. Although different forms of evolutionary theory assign varying degrees of importance to natural selection, they all suggest that the complexity found in living things need not have developed at once, but could have been acquired piecemeal. Proponents of selectionist approaches to evolution emphasize the claim that each potential step in the acquisition of complexity is tested by natural selection.1 It is tacitly supposed that each increment of change towards an eventual complex structure would be of benefit to the organism that bears it, and would therefore be preserved by natural selection. Though in no way goal-directed, the outcome of this process, repeated often enough and long enough, would be a complex living system.
Biochemist Michael J. Behe,2 though an evolutionist, has challenged this widely held and incessantly taught notion:
‘What type of biological system could not be formed by “numerous, successive, slight modifications”? Well, for starters, a system that is irreducibly complex. By irreducibly complex, I mean a single system composed of several well-matched, interacting parts that contribute to the basic function, wherein the removal of any one of the parts causes the system to effectively cease functioning. An irreducibly complex system cannot be produced directly (that is, by continuously improving the initial function, which continues to work by the same mechanism) by slight, successive modifications of a precursor system, because any precursor to an irreducibly complex system that is missing a part is by definition non-functional’ [italics in original].
Behe then presents several biochemical examples of irreducible complexity. Certain evolutionists, notably those reviewing Behe’s book, summarily dismiss his argument and insist, in a purely arm-waving manner, that there is no such thing as an irreducibly complex system. Given enough time, with the wonder-working power of natural selection acting upon genetic mutations, even seemingly impossible things can happen. Yet despite this bravado, there are other evolutionists who, without mentioning Behe or using the phrase irreducible complexity, acknowledge that it is a very serious problem for evolutionary theory. A few such examples, focusing on gene regulation and novel protein origins, are presented here.
Origin of genomic regulatory systems
The nature and degree of gene expression is commonly governed by a tightly intertwined set of regulatory elements found on the DNA molecule. How is such a concert of regulatory elements supposed to have evolved as a unit? The scenario invoked has a distinct flavour of storytelling:
‘Advantageous substitutions in regulatory elements caused by genetic factors are most interesting. They must be responsible for morphological evolution as discussed before. When a new chain of gene expression patterns for transcription factors and signal transduction elements is appearing, many advantageous mutations are thought to occur simultaneously at the loci participating in the chain. This process is called “recruitment” or “cooption” by developmental biologists.
How such a chain originates is a very difficult problem, i.e., a module of interacting gene loci would have to be constantly tested by natural selection under various genetic and external factors. On very rare occasions, while wandering via mutation and drift under available transcription factors, a module might find its place in a larger gene regulation network. Then positive selection may work on the regulatory elements of the module loci’3 [emphasis added].
Yes, and if a cow had wings, it might fly. Various speculative attempts to overcome the problem of irreducible complexity, discussed in the remainder of this report, are all essentially hoped-for simultaneous accidental changes in the genome that are supposed to eventually lead to the emergence of biological novelty. In the past, these have been called hopeful monsters. However, owing to the fact that the mechanisms discussed are not as extreme as the classical hopeful monsters,4 I informally refer to them as mini hopeful monsters.
A network of highly regulated genes governs the development of an organism. One of the chief difficulties of overcoming the irreducibly complex system of such gene regulation is the fact that, not only do all the parts of such a system interact closely, but it is, except under special circumstances, difficult to upset this finely-tuned system:
‘Evolution exploits genetic differences between individuals in order to remodel developmental programs, yet development is generally robust to individual genetic differences and environmental perturbations. Theoretical models describe how developmental homeostasis is developed and why it is maintained, as well as how it could be disrupted so that evolutionary change can occur.’5
In the common fruit fly, Drosophila, environmentally mediated perturbations of the Hsp90 gene can cause the simultaneous deregulation of a number or genes. This, in turn, causes these flies to display a variety of developmental abnormalities, such as deformed or absent eyes, notched wings, duplicated bristles, etc.6 Such malformations hardly inspire confidence in this mechanism as a cause of alleged evolutionary change.
This is not to say that simultaneous changes, which result in the uncovering of biologically meaningful cryptic genetic variation, cannot occur in the genome. In fact, several examples are reviewed, by this author,7 in conjunction with the appearance of useful ‘hidden’ variation among the numerically impoverished organisms of the post-Flood world. However, note the minuscule scale of these changes. Clearly, disruptions of gene complexes are a necessary but not sufficient cause for the appearance of new irreducibly complex structures. The simultaneous appearance of several mutations, even if neutral or beneficial, is not yet proof that any combination of them can produce even one new irreducibly complex system!
Notice, in the following quote, the huge leap between potential deregulation of gene complexes, on one hand, and the hoped-for emergence of major evolutionary changes as an eventual outcome of this mini hopeful monster mechanism:
‘By altering the activities of multiple signal transducers and thereby simultaneously weakening several developmental pathways, Hsp90 can expose such variation, allowing selection to remodel many different processes at once … . The use of Hsp90 as a capacitor for the conditional release of stores of hidden morphogenic variation may have been adaptive for particular lineages, perhaps allowing the rapid morphological radiations that are found in the fossil record.’8
The emergence of new biological functions
There is no doubt that irreducible complexity is acknowledged in the following statement:
‘A major enigma in evolutionary biology is that new forms or functions often require the concerted efforts of several independent genetic changes. It is unclear how such changes might accumulate when they are likely to be deleterious individually and be lost by selective pressure’9 [emphasis added].
One hopeful mini monster mechanism, proposed three decades ago by Koch,10 postulated that genes could be temporarily inactivated, allowing them to drift neutrally (acquire a variety of random mutations without the possibility of being removed by natural selection). Subsequently, the genes would re-acquire function, and the multiple accumulated mutations could then be tested simultaneously by natural selection. Eventually, a lucky combination of mutations would turn out to be beneficial to the host organism, and even cause the sudden appearance of biological novelty. It is now recognized that this postulated inactivation-reactivation process is not likely to be effective:
‘However, the known mechanisms for the reactivation of inactive genes work sporadically, act infrequently and provide no obvious means for sampling coding changes in several genes simultaneously.’11
True and Lindquist12 propose an alternative scenario for the accumulation of temporarily neutral mutations. It is based on a prion (proteinaceous infectious particle)-mediated mechanism that modulates the effectiveness of yeast genes’ premature stop codons in the termination of transcription, thus allowing for flexibility in terms of gene expression. It is suggested that, while the premature stop codon is ‘on’, mutations occurring in the gene sequence beyond this codon can accumulate freely owing to the inability of this part of the gene from being ‘read’, and therefore potentially impacting host fitness. But once this prion [PSI+] turns the premature stop codon ‘off’, the previously ‘unreadable’ part of the gene sequence becomes expressed and the phenotypic effects of the accumulated mutations can then be simultaneously tested by natural selection.
This particular mechanism has been shown to cause changes in such things as the heat resistance of the affected yeast, the ability of the yeast to grow in different chemical media, and the geometric shape of the yeast colony itself.13 Although new phenotypes appear to have been produced (or at least unmasked) by this particular mechanism, their impact is rather modest. The new phenotypes appear to be little more than the ‘tweaking’ of pre-existing yeast traits rather than the emergence of radically new yeast behaviors or capabilities. In any case, the yeast is still yeast. How are the cumulative effects of this mechanism supposed to add up to the emergence of irreducibly complex structures? Consider the following:
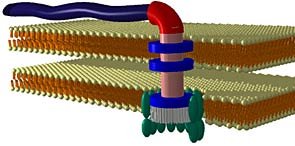
The bacterial flagellum. In an attempt to side-step irreducibly complexity, evolutionists hypothesize that the components of irreducibly complex systems all had other functions before being co-opted for use. These prior functions are largely speculative. As an ever increasing number of irreducibly complex systems are discovered, and more and more components in each system, the number of ‘functions’ (and thereby stories) needing to be found is ever increasing.
‘How might such a system evolve and be maintained? We suggest three different, not exclusive possibilities … The phenotypic diversity generated by these ORFs [gene’s open reading frames, subject to having their premature stop codons turned off and then on] by [PSI+] would depend upon which ORFs had acquired ISCMs [inactivating stop-codon mutations], the frequency of their readthrough, and the presence of additional mutations they have acquired while in the inactive state … Such mechanisms may be present more broadly than previously suspected and exert an important influence on the rates and mechanisms of evolutionary change’14 [emphasis added].
Again, the foregoing has the distinctive flavour of storytelling. In any case, we once again see a huge leap in reasoning from the observed very small phenotypic changes all the way to the hoped-for emergence of totally new structures and functions.
Origin of new proteins: still more hopeful mini monsters
Earlier, the old ideas of Koch15 were discussed in conjunction with the evolutionists’ difficulty of accounting for simultaneous large-scale beneficial changes cumulating in the appearance of new biological functions. When it comes to the supposed origin of new proteins from pre-existing ones, a step-by-step set of changes likewise appears to be untenable. A protein having an intermediate sequence between that of its ancestral form and its eventual descendant form would likely be deleterious to its host (if able to be translated at all), and so would not be preserved by natural selection in the first place. In fact, Koch16 had recognized this fact, and so had aptly titled his paper: The importance of untranslatable intermediates. This, in fact, is the central problem for all evolutionary attempts to explain the origin of all irreducibly complex structures.
Using recent thinking and vocabulary, Harrison and Gerstein,17 having tacitly recognized the irreducible complexity of protein design, have attempted to update Koch’s old hypothesis:
‘How does one get unique folds in certain phylogenetic groups? As shown in Figure 5(b) [see next paragraph], in some cases it may be difficult to imagine a scenario for this where each intermediate form has to be a functioning protein that is transcribed and translated. (This is in contrast to other evolutionary pathways, where functioning and selected intermediates are more plausible). One can speculate that resurrectable pseudogenes could eliminate this paradox to some degree. A sequence comprising a particular domain fold or (more likely) part of a domain could become pseudogenic. It could then drift freely as a pseudogene, and evolve to a new domain fold upon or after resurrection. In this scheme, each intermediate does not have the constraint that it be a folded functional protein’ [emphasis added].
The word speculation, used in the quote above, is an excellent choice of words for this hopeful mini monster mechanism! And, as in all prior discussed instances, speculative outcomes do not begin to explain the origin of irreducibly complex proteins at all. (Note that Figure 5b in the quote above simply shows changes in geometric shapes to illustrate the (virtually impossible) gradual change in proteins vs the hopeful accumulation of this fortuitously beneficial set of changes within nonfunctional pseudogenes that will one day again become functional genes). Although there are claims18 about supposedly nonfunctional gene copies (pseudogenes) becoming eventually resurrected as new functional genes, such instances are few and far between (not to mention the fact that all inferences of genes changing to pseudogenes and back to genes rely on phylogenic analyses and thereby presuppose organic evolution). This returns the evolutionist to the problem of the rarity of this presumed phenomenon, as already shown by the earlier quote from True and Lindquist.19
Other attempts at understanding the hoped-for evolutionary origin of de novo proteins have also been undertaken. For instance, Taverna and Goldstein20 noted the fact that proteins found in living systems, in contrast to synthetic proteins, retain their structure, stability, and function even in the face of a significant number of alterations in sequence. Citing and then extending some theoretical experiments revolving around evolution of individuals as part of a group, they suggest that proteins found in living things have this capability because they evolved that way:
‘Why does the sequence plasticity observed in site-directed mutagenesis not translate into ease in protein engineering? … These results suggest that the observed sequence plasticity of biological proteins may occur because these proteins have evolved to be robust to these specific experiments … Firstly, the lessons of sequence plasticity in biological proteins may be inapplicable to artificially designed proteins. It may be necessary to have a de novo sequence exquisitely designed to have properties similar to biological proteins.’
This consideration begs the question about the very origin and diversification of proteins in the first place!
Discussion and conclusions
I have discussed only a few examples of irreducible complexity that came to my attention inadvertently (while researching other topics). For this reason, no inferences should be drawn regarding the extent of irreducible complexity based on this short report.
Are they the best of both worlds or are they the worst of both worlds?
The traditional conception of step-by-step major evolutionary change has the supposed advantage of reasonable probability for each step while suffering from the disadvantage of being incapable of producing the necessarily simultaneous changes (hence irreducible complexity). Hopeful monster scenarios reverse this situation, invoking a very improbable event to (theoretically) giving rise to a simultaneously emplaced set of interconnected simultaneous changes. The mini hopeful monster scenarios discussed are intermediate between the foregoing two approaches to the understanding of alleged major evolutionary change. But are they the best of both worlds or are they the worst of both worlds?
Consider the central fact that all the changes discussed in the cited works are quite small. Accounting for new irreducibly complex structures by the foregoing mechanisms is a completely different proposition. There is not the slightest indication, much less proof, that such changes (or ones comparable to those discussed in the cited works) could ever accumulate in a manner that eventually produces a totally different life form (i.e. commensurate with a higher-level taxonomic category). It seems clear that a succession of mini hopeful monster events, capable of originating a de novo irreducibly complex system, appears to be simultaneously improbable and incapable of effecting the large-scale simultaneous changes. Using Behe’s analogy of the mousetrap,21 one mini hopeful mini monster event may perhaps theoretically happen to produce a hammer that could fit with other would-be mousetrap components. Yet there is not the slightest indication that successive hopeful mini-monster events would also produce the requisite correctly shaped and correctly deployed spring, catch, holding bar, etc.
It almost seems as though evolutionists are invoking these hopeful mini monster mechanisms as an act of desperation. In any case, the giant chasm that remains between the observed tiny changes, on the one hand, and the speculated large-scale evolutionary outcomes, on the other, itself attests to the validity and force of the argument of irreducible complexity.
Footnotes
- Dawkins, R., The Blind Watchmaker, Norton and Co., New York, 1986.
- Behe, M.J., Darwin’s Black Box, Free Press, New York, p. 39, 1996.
- Ohta, T., Near-neutrality in evolution of genes and gene regulation, Proceedings of the National Academy of Sciences of the USA 99(25):16134–16137, 10 December 2002, p. 16136.
- The classical example of a hopeful monster would be a fully-fledged bird, which hatches from the egg of a reptile.
- Rutherford, S.L. and Lindquist, S., Hsp90 as a capacitor for morphological evolution, Nature 396(6709):336–342, 26 November 1998, p. 341.
- Rutherford and Lindquist, Ref. 5, pp. 336–340.
- Woodmorappe, J., Noah’s Ark: A Feasibility Study, Institute for Creation Research, El Cajon, pp. 189–190, 1996. Such small-scale simultaneous changes would be useful for such things as the ecological differentiation of post-Flood organisms, but not for the appearance of new irreducibly-complex structures.
- Rutherford, S.L. and Lindquist, S., Hsp90 as a capacitor for morphological evolution, Nature 396(6709):336–342, 26 November 1998, p. 341.
- True, H.L. and Lindquist, S.L., A yeast prion provides a mechanism for genetic variation and phenotypic diversity, Nature 407(6803):477–483, 28 September 2000, p. 477.
- Koch, A.L., Enzyme evolution: I. The importance of untranslatable intermediates, Genetics 72(2):297–316, October 1972.
- True, H.L. and Lindquist, S.L., A yeast prion provides a mechanism for genetic variation and phenotypic diversity, Nature 407(6803):477–483, 28 September 2000, p. 477.
- True and Lindquist, Ref. 8, p. 473–483. See especially p. 482.
- True and Lindquist, Ref. 8, p. 473–483. See especially p. 482.
- True and Lindquist, Ref. 8, p. 473–483. See especially p. 482.
- Koch, A.L., Enzyme evolution: I. The importance of untranslatable intermediates, Genetics 72(2):297–316, October 1972.
- Koch, A.L., Enzyme evolution: I. The importance of untranslatable intermediates, Genetics 72(2):297–316, October 1972.
- Harrison, P.M. and Gerstein, M., Studying genomes through the aeons: protein families, pseudogenes and proteome evolution, Journal of Molecular Biology 318(5):1155–1174, 17 May 2002, p. 1170.
- Harrison, P.M. and Gerstein, M., Studying genomes through the aeons: protein families, pseudogenes and proteome evolution, Journal of Molecular Biology 318(5):1155–1174, 17 May 2002, p. 1170.
- True, H.L. and Lindquist, S.L., A yeast prion provides a mechanism for genetic variation and phenotypic diversity, Nature 407(6803):477–483, 28 September 2000, p. 477.
- Taverna, D.M. and Goldstein, R.A., Why are proteins so robust to site mutations? Journal of Molecular Biology 315(3):479–484, 18 January 2002.
- Behe, Ref. 2, pp. 42–43.
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis