

The Intricacies of Flight in the Natural World
Examples of complexity in the natural world are not hard to come by. Living creatures all are examples of irreducible complexity. This phenomenon is well known in design engineering and refers to the fact that there are mechanisms which only work when everything works together. This is certainly true in the natural world as many mechanisms right down to the molecular world show that this is the case. The cell will not work without all the DNA machinery being in place, as very ably demonstrated by Behe.1 One of the best examples of complexity which defies a series of “gradual” changes is flight. Dawkins2 sought to try to justify such an idea of gradually producing flight, but showed very little understanding of the fundamental engineering principles involved in aerodynamics and control. He even suggested there may be wingless ancestors that, living in water, raised their gills to make primitive sails which then evolved into the flapping wings of insects as they got lifted by the wind.3 Insect flight and flapping motion is far more than having appendages to the body, and all engineers know this.
For controlled, heavier-than-air flight, there are four fundamental requirements: (1) A correct wing shape to give a lower air pressure on the upper surface; (2) a large enough wing area to support the weight; (3) some means of propulsion or gliding; and (4) extra surfaces, or a means of altering the main surfaces, in order to change direction and speed.
Flight occurs in many groups of the living world: (a) birds; (b) insects—flies, bees, wasps, butterflies, moths; (c) mammals—bats; (d) reptiles—the extinct pterosaurs (e.g., pterodactyls and pteranodons).
In reality each class of creature in the supposed development of each case of flight is in fact anatomically different with no connection made even by the most ardent evolutionist.
In each case the wings are substantially different and there is no evidence whatsoever of any connection between the supposed evolutionary development of any of these creatures. The evolutionist is faced with not just one impossible hurdle—that some reptiles grew feathers and began to fly—but two other hurdles. These are that flight evolved again when some rodents (mice? shrews?) developed a skin-like surface and developed into bats, and then quite separately some insects grew very thin scales to become flies, bees, and butterflies. The theory is called convergence in evolutionary literature. However, in reality each class of creature in the supposed development of each case of flight is in fact anatomically different with no connection made even by the most ardent evolutionist. The detail of the proposition that in each case there was a transition leading to the emergence of flight is where the so-called convergence position becomes untenable. For birds the main proposal has been that small reptiles (dinosaurs) led to birds. It is seriously proposed that there was a “pro-avis” dinosaur that flapped scales on its arms to catch insects, and then changed its scales to feathers to gain airborne advantage on its prey. For bats it is proposed that a shrew or mouse developed wings, and for insects there has been no coherent theory presented for the origin of their wing structures—in many ways that is the hardest of all, since flying insect fossils are found deep in the Paleozoic strata.
Even if one accepts the fossil record as a guide to change over millions of years, there is no firm evidence of any “pro-avis” creature ever existing in the fossil record, there is no evidence at all for an intermediate bat wing structure, and flying insect fossils indicate no precursors whatsoever.
So for the evolutionist, there is the scenario of flight evolving at least three times independently with no fossil evidence to support such a notion. The wings of the three main current groups of flying creatures today are substantially different—birds wings are made of feathers, insect wings of scales, and bat wings use skin spread out over a skeleton.
Birds
Feathers
A bird’s wings are made of feathers. A feather is a marvel of lightweight engineering. Though light, it is very wind-resistant. This is because there is a clever system of barbs and barbules with hooks on one side and ridged barbules on the other. Thus an ingenious mechanism exists for keeping the surface flexible and yet intact. Each barb of a feather is visible with the naked eye and comes off the main stem. What is not generally realized is that on either side of the barb are further tiny barbules which can only be seen under a microscope. These are of a different type depending on whether they are coming from one side of the barb or the other. On one side of the barb ridged barbules will emerge, while on the other side, the barbules will have hooks on. Thus the hooks coming out of one barb will connect with ridges reaching in the opposite direction from a neighboring barb. The hooks and ridges act like Velcro, but go one stage further, since the ridges allow a sliding joint, and there is thus an ingenious mechanism for keeping the surface flexible and yet intact. The next time you see a flight feather on the ground, remember it is a marvel of lightweight flexible aerodynamic engineering. Reptile scales have no hint of such complicated machinery. Stahl has freely admitted,
No fossil structure transitional between scale and feather is known, and recent investigators are unwilling to found a theory on pure speculation.4
There is no genetic information within reptile scales to allow such a unique device as the sliding joint of a feather to be made. The tortuous route suggested by some of small “advantaged mutations” to scales leads to clumsy structures which are in fact a disadvantage to the creature. Not until all the hook and ridge structure is in place is there any advantage, even as a vane for catching insects! Unless one invokes some “thinking ahead” planning, there is no way that chance mutations could produce the “idea” of the cross-linking of the barbules to make a connecting lattice. Even if the chance mutation of a ridge/hook occurs in two of the barbules, there is no mechanism for translating this “advantage” to the rest of the structure. This is a classic case of irreducible complexity which is not consistent with slow evolutionary changes, but quite consistent with the notion of design.
Unless one invokes some “thinking ahead” planning, there is no way that chance mutations could produce the “idea” of the cross-linking of the barbules to make a connecting lattice.
But that is not all. Even if one had the feather, the delicate lattice structure would soon become frayed unless there was also oil to lubricate the sliding joint made by the hooked and ridged barbules (see Fig. 1). Most of us realize that once the barbs of a feather have been separated, it is difficult to make them come back together. The feather becomes easily frayed in the absence of oil, which a bird provides from its preening gland at the base of its spine. Some of this oil is put on its beak and spread throughout the feathers, which for water birds also gives waterproofing of its surface (thus water slides off a duck’s back). Without the oil the feathers are useless, so even if a supposed land-dwelling dinosaur got as far as wafting a wing, it would be no use after a few hours!
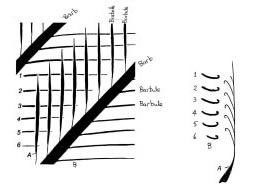

Figure 1. Detail of hook and ridge system for pennaceous feather barbules. Photograph courtesy of Prof. David Menton, Washington University School of Medicine, St. Louis, Missouri.
Bone Structure
As one might expect, however, the story does not end there either, for a bird can only fly because it also has an exceedingly light bone structure which is achieved by the bones being hollow—all except some diving birds like the loon of Canada (like a duck but with a pointed beak), which has some solid bones for diving. All birds other than some diving birds such as Divers (called loons in America) and puffins, for example, maintain the skeletal strength by cross members within hollow bones. Diving birds have some solid bones to aid the dive into the water. The arrangement of hollow structures began to be used in the 1930s and 40s for aircraft wing construction and is termed the Warren truss arrangement. Large birds such as an eagle or a vulture would simply break into pieces in mid-air, if there was some supposed halfway stage in their skeletal development where they had not yet “developed” such cross members in their bones.
Muscle Arrangement
Consider the wing-flapping motion of a bird. The wing-flapping motion of a bird is operated by two muscles operating in tandem. One is called the pectoralis major muscle, which runs across the chest (in a similar way to mammals and reptiles) and for a bird is attached to the base (keel) of the breast bone (sternum). However, the other muscle is unique to birds. For mammals and reptiles the raising of arm (that is the humerus bone) is achieved by contracting a muscle in the back. However, a bird uses a supracoracoideus muscle, which is attached to the keeled sternum at one end and at the other is threaded through a hole in the coracoid bone and attached from above to the humerus bone. Consequently, contraction leads to a pulley effect and the lifting of the wing in the upstroke. So flapping motion (particularly taking off from stationary) is achieved by the powerful operation of these two muscles in tandem. To suggest that the muscles evolved alongside a unique skeletal arrangement (involving an enlarged keeled sternum with a special hole in the coracoid bone) is imagination and not the careful deduction from empirical science.
Breathing Mechanism of Birds
Furthermore, birds breathe differently, as indicated in Fig. 2. Due to the necessity for birds to operate at high energy exchange rates with the muscles of their wings (in particular), they need a high source of energy intake to operate them, so that normal respiration is not good enough. Human beings breathe about 12 times per minute whereas small birds breathe about 250 times per minute. To achieve this, the oxygen-rich air is fed straight into air sacs which are then connected directly to the heart, lungs, and stomach.
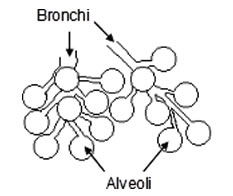
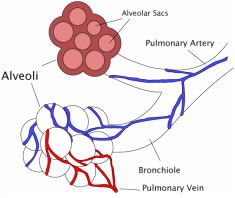
Figure 2. The end flow mass exchanger in mammalian and reptilian respiration.
A remarkable feature is then crucially different in the avian lung arrangement. Unlike the mammalian and reptilian requirement where air containing carbon dioxide must be breathed out first before the next intake of oxygen, in the avian system the oxygen-rich air is passed to a rear air sac, and is then continuously fed through the lung (see Fig. 3) from a rear air sac through parabronchi tubes in the lung to a forward air sac. Consequently, the air never stops as in a reptilian or mammalian system. Remarkably, it takes two inhalations and exhalations for one complete cycle to be achieved. The rate is also very different. Human beings breathe about 12 times a minute, whereas small birds can breathe up to about 250 times a minute. This is thus a perfect system for the high metabolic rate in birds, which use up energy very quickly. Furthermore a bird’s breathing system is operated by the sternum (breast) bone and it has no diaphragm muscle. Any claimed in-between transition would have not been able to breathe as one system changed into another. Note also that birds are warm blooded, which presents a vast biological hurdle for those who maintain a reptile ancestry for birds (since almost all reptiles are cold blooded).
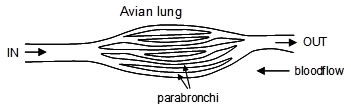
Figure 3. Idealized schematic of parallel counterflow mass exchange system in avian respiration.
Integrated Design of Birds

Figure 4. Male American Kestrel. Photo courtesy of R. W. Scott.
Birds then require the integration of many features. In the flapping motion a bird requires the unique pair of muscles, with a forward facing elbow joint to enable the foreshortening of the wing used much in the upward stroke of most species, and in the dive of birds of prey. The versatility of the swivel joint at the base of the wing, coupled with the elbow joint on the wing itself and the smooth feather structure overlaying all, leads to great flexibility in the aerodynamics of the wing. Lift and drag can be balanced with instant movements, which in aircraft still require comparatively cumbersome changes of flap and ailerons. Suppose we have an “almost” bird with all the above structures—such as feathers, preening gland, hollow bones, the unique avian lung, warm blood, enlarged keeled sternum bone, a hole in the coracoid bone, a swivel joint and forward facing elbow joint—but no tail! Controlled flight would still be impossible. Longitudinal stability can only be achieved with a tail structure, which most small boys soon realize when making paper airplanes! But what possible advantages do all of the above have for any land-based “almost” bird? Such a creature would be easy prey to any hunting animal. In the list of mechanisms (feathers, preening gland, and so on), all are essential. One can attempt to drop one and the whole project fails! The tail is essential (as anyone will have realized in observing a kestrel hovering by flipping the outer portion of its wings and balancing against its tail—see Fig. 4), and with the tail must come another muscle to operate the variable small, but all-important wing surface—for instance, holding the plumage spread out and downwards when coming in to land. In other words the tail is no use as a static “add-on.” It must have the means of altering its shape in flight. All these mechanisms are controlled by a nervous system connected to the onboard computer in the bird’s brain, all pre-programmed to operate within a wide envelope of complicated aerodynamic maneuvers.
For birds to have evolved from reptiles, all the features—hollow bones, feathers, unique avian lung, twin muscle arrangement, enlarged keeled sternum bone with hole in coracoid, and tail—must all develop simultaneously. Such a notion is not consistent with careful scientific observation, which points to the obvious alternative that this basic system is the product of design.
Humming Birds
One of the most delightful demonstrations of all of the above principles coming together is with the humming bird. These small birds have the ability to beat their wings at up to 80 beats per second, and as is well known, can hover, fly backwards, forwards and sideways with ease (much of the information here is drawn from an excellent article by Denis Dreves5). Speeds of 50 miles an hour are commonplace for these flying marvels. Fuel must be replenished very quickly because of the great turnover of energy. Consequently it must feed on a food which can be broken quickly down into energy. All this is achieved by these birds feeding on the nectar of flowers, requiring the ability to hover and the thin, long beak to get into the flower (e.g., a fuschia for the rufous humming bird). It also has a special tongue which has two furrows enabling the nectar to be stored on the tongue. The long tongue goes in and out of the bill at an unbelievable rate of 13 times per second, and when retracted is curled up at the back of the head. One can envisage the odd scenario of the supposed half-evolved humming bird either with the ability to hover and a sparrow beak, unable to feed, or the long beak, but no ability to hover, flying into the flower, but with no ability to stop. All the requirements must be there to begin with. The extreme maneuverability of humming birds is due to their having the ability to swivel the wing through a much greater angle than for other birds. Consequently, the humming bird can produce a power stroke on the upward motion of the wing as well as the down stroke, and the motion of the wing tip of a humming bird in flight sweeps out a figure of eight as the joint swivels round first some 90 degrees in one direction, and about 90 degrees in the other direction. Further rotation is possible, which means that the wing can thus beat a power stroke in any direction, with small asymmetries enabling sideways movements as well.
Transitional Fossils?
The flight of birds cannot be explained by supposed evolutionary change. The attempts to find any transitional forms have all failed. Archaeopteryx has been shown to have fully-developed flight feathers (thus no half-bird) with other recognizable birds found fossilized at a lower level. Other supposed “pro-avis” creatures (half reptile/half bird) have never been found, though some imaginative attempts have been made from the Liaoning shales in China. All these creatures are either extinct birds or have a fur which indicates they were possibly extinct mammals. None show any evidence of half feathers. The evidence taken as a whole is overwhelming that birds have always been birds, and is entirely consistent with their being created right at the beginning on Day Five just as the Bible says. It is not scientific to argue on the one hand for the obvious design of a Boeing 747, and then rule design “out of court” when considering the far more versatile flight of an eagle, falcon, or the remarkable hummingbird. Modern minds within the secular media are presenting an unscientific duality of thought when considering on the one hand engineering complexity in man-made machines, glorying in the great creative advances of mankind, and yet on the other hand presenting the complexity in the world around us (of often far greater intricacy than man-made machines) as due to a gigantic, unplanned cosmic experiment with no Creator.
Flying Insects
Flying insects have no connection whatsoever with birds, and yet flight is fully developed in all the fossils of flies, bees, moths, and butterflies.
Flying insects have no connection whatsoever with birds, and yet flight is fully developed in all the fossils of flies, bees, moths, and butterflies. There is not any contender at all for any transitional forms. The wings of such creatures are exceedingly fragile, made of scales, membranes, or sometimes hairs or bristles. They start as larvae or grubs, with some feeding on totally different substances than the adult (e.g., caterpillars and butterflies). The best example is the dragonfly, which starts as a nymph underwater, obtaining oxygen from the water, yet no adult dragonfly could possibly exist in the same surroundings. It is quite common for dragonflies to reach speeds of 30 miles per hour, but no transitional fossils exist. The evidence is in fact of much larger fully developed dragonflies6 with a wingspan of two to three feet existing in the past, indicating decline today, rather than advance. The related damselfly has the remarkable ability to hover as well, such that the sophisticated aerodynamics of the four wings operating in an asymmetric fashion inspired the design of early forerunners of the modern helicopter.
The complexity of the life cycle of creatures such as the butterfly (caterpillar to chrysalis and then to butterfly) and the dragonfly (water nymph to dragonfly), and their perfect wing structure as adults, points to intricate design which cannot be explained by small changes. The survival of each species is dependent on all the mechanisms being present to begin with. The second body plan of the adult butterfly/dragonfly is absolutely essential for the next generation to start, as caterpillars/water nymphs do not lay eggs. This amazing feature of these creatures (called metamorphosis) must have been there right from the beginning of these creatures existing. And the rare fossils found of butterflies, dragonflies and damselflies confirms this.
Migrating Butterflies

Figure 5. The Monarch Butterfly lays its eggs on milkweed, which is poisonous to most predators of larvae.
The Monarch butterfly of North America (Fig. 5) migrates 2000 miles from California in the northwest (or Ontario in the northeast) to the over-wintering site of Central Mexico. More amazing is the following fact: some adults who make the journey back are fully mature, so that some females lay eggs and die en route northward. These offspring after going through the caterpillar/chrysalis stage then continue the migration northward. More remarkable still is that not all these make it back to northeastern America and another third generation then finally completes the journey, fulfilling the aspiration of grandma! This of course means that a remarkable system of information is bound up in the genetic coding of each butterfly, such that it “knows” at what stage the group of butterflies is in the migrating cycle—this information is passed on to each generation. Such a delicate mechanism shouts intelligent design.
Furthermore, it has been established that magnetite has been found in the bodies of monarch butterflies (and also honeybees), indicating that they are able to orientate themselves by sensing the earth’s magnetic field. Their eyes are also sensitive to polarized light from the sun, again giving them a direction cue. The two eyes, far from being simple, are actually each made of 6,000 separate lenses! There are no half-formed butterflies in the fossil record, and they are similar to modern ones—fully formed.
Flying Mammals—Bats
Bats are entirely different again from birds and insects, with wings made of skin, and a sonar system on some bats enabling echo sounding with pinpoint accuracy, and thus enabling the bat to find insect food on the wing, with unbelievable precision. No half-bat has ever been found in the fossil record, and it would be hard to imagine how such a halfway creature could survive (see Fig. 6). Flight alone is only possible with fully-developed wings. This combined with the sophisticated sonar is yet another example of irreducible complexity.
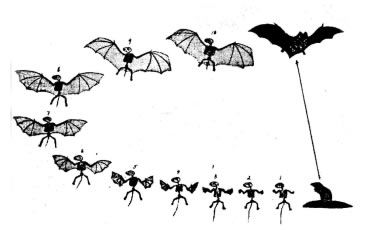
Figure 6 Supposed transitional changes for bat evolution—none exist in the fossil record save the shrew and the bat.
Conclusions
As a scientist, I see nothing to discount straightforward belief in Scripture, when considering the mechanisms in nature of which flight is but one part. Those which have been considered in relation to flight could be added to by many more intricately balanced mechanisms which overwhelmingly testify to a Creative hand.
Many of course refuse to acknowledge the evidence for design in nature because they make the untestable assumption of atheism. If someone seriously did doubt the design of a modern airliner, that person could be convinced by taking him into an aircraft factory and introducing him to the teams of design engineers. In the same way, man’s prejudice against design in creation can only really be answered by a radical change of heart and by meeting the Author of all personally. In the end the difference between these two worldviews is due to religious differences. It is because men do not want to be accountable to a Creator God (Romans 1:18) that they persist with a theory which has little evidence to support it.



Footnotes
- Michael Behe, Darwin’s Black Box: The Biochemical Challenge to Evolution (New York: Free Press, 2006).
- Richard Dawkins, Climbing Mount Improbable (New York: Viking, 1996), 97–125.
- Ibid., note on p. 125.
- Barbara Stahl, Vertebrate History: Problems in Evolution (New York: McGraw-Hill, 1974), 349.
- Denis Dreves, “The Hummingbird: God's Tiny Miracle,” Creation Ex Nihilo 14, no. 1 (1992): 10–12.
- David Attenborough, Discovering Life on Earth (Boston: Little, Brown & Company, 1988), 60–61.
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis