

Image from Answers magazine
Cranial Capacity and Endocranial Casts
Originally published in Journal of Creation 4, no 1 (April 1990): 56-65.
Abstract
Man has always prided himself on his intellectual capabilities and has often pondered to explain the reason why he alone is able, unlike the rest of the animal kingdom, to contemplate and communicate.
Introduction
Man has always prided himself on his intellectual capabilities and has often pondered to explain the reason why he alone is able, unlike the rest of the animal kingdom, to contemplate and communicate with others. Since man’s brain is larger than other animals, it is natural for man to conclude that the brain is the hallmark of man and the measurement of it must be the key to the understanding of his unique intellectual capacity.
Measuring of man’s potential intellectual capacity from sources other than direct observations and testing is derived from direct, indirect and inferential data. The direct data is obtained from measuring cranial capacity and examining casts which ‘expose’ the interior of the skull. The indirect evidence comes from the study of comparative brain sizes, which is based on the assumption that there is a correlation between brain size and configuration, and behavioural studies. Other indirect evidence comes from the study of fossil remains of non-cranial parts which infer manual dexterity and visual acuity.
The inferential evidence, although conjectured, is important in measuring potential intellectual capacity. The main evidence for complexity of behaviour is deduced from tools and indications for ‘cultural norms’ such as hunting organizations, religious implications, etc.
This paper will expose the myth that man’s potential intellectual capacity can be measured from his cranial capacity and configuration.
Measuring cranial capacity
Cranial capacity has been measured in a variety of ways. Mustard, millet or flax seeds are used because they closely approach the volume equivalent of a liquid, but also small lead shots are commonly employed.1If mustard seeds are used, they are placed into the hollow skull, after the small foramina are plugged with cotton wool through the foramen magnum, by means of a funnel and agitated by the hand or an agitating instrument. When the skull is sufficiently packed, the material is poured into a measuring glass which is again agitated and packed. Different measurements on the same specimen can be made depending upon the way seeds are compressed, the speed with which the seeds are shot in, and the diameter of the neck of the funnel.
A more accurate way of measuring cranial capacity, though rarely done, is to make an endocranial cast and measure the amount of water the cast displaces. Holloway2 has shown that cranial capacity estimates of many major hominid fossils differ when measured by the other methods.
Indirect measurements for cranial capacity can be made with different formulae depending upon sex. Olivier3 lists these formulae:
Male: 359.34cm3 + 365 x 10-6 (Length x breadth x auricular height)
Female: 296.4cm3 + 375 x 10-6 (Length x breadth x auricular height)
Changes are made in formulae depending upon the race and the thickness of the parietal bone.
Often fossil hominids are found fragmented and are reconstructed upon a paradigm according to the law of correlation. Jerrison4 noted the paucity of whole crania of australopithecines and habilines and remarked that cranial statistics seem to conform to foci of averages based on a few reconstructions.
Because man’s cranial capacity is so variable today, it has been shown that there is very little relationship between cranial capacity and human intelligence. Various populations are found to have capacities within the ‘fossil man’ range. Textbooks show the assumed evolution of man’s brain (see Figure 1). However, a Melanesian with a cranial capacity of 790 cubic centimetres (cc) is said to be the lowest on record of a normal adult.5 Harris6 claimed that the variability of man’s cranial capacity starts at 850cc. Although mentally retarded adults have had measured cranial capacities of 511cc and 519cc which are equal to an adult gorilla’s, their behaviour was obviously not pongid.
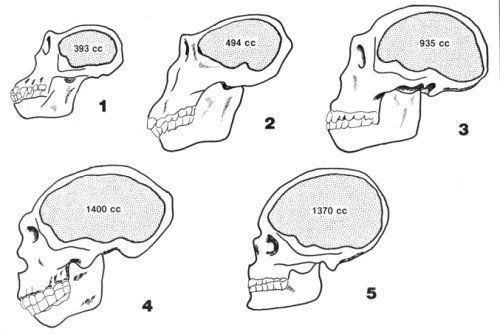
Figure 1. A typical illustration in textbooks showing the supposed evolution of man’s cranial capacity. 1. Chimpanzee 2. Australopithecine 3. Homo erectus 4. Neanderthal 5. Cro-Magnon
Therefore, as Holloway7 succinctly remarks:
‘One cc of chimpanzee cortex is not equivalent of human cortex, nor is it likely that any equivalent measure can be found.’
So it is evident that modern man overlaps the Homo erectus range of cranial capacity which is 800 to possibly 1400cc. (Note the upper range applies to the Vertezollos occipital bone.)
The usefulness of cranial capacity measurements to the assessment of potential human intelligence is criticized by Weidenreich8 who questions:
‘Is it possible to infer from the size of the brain the degree of intelligence and cultural efficiency of its bearer, regardless of whether this bearer lived several hundred thousand years ago or lives today? Almost everyone, layman as well as scientist, seems to be convinced that such a correlation is a well established fact … . However, we do not know of any fact which proves that the mere increase of the size of the brain is tantamount to an advance in mental ability.’
Clark9 makes this supporting statement.
‘… So far as it has been possible to apply appropriate tests, there is within such limits no marked correlation between the brain size and intelligence. To the paleoanthropologist this lack of correlation is particularly disconcerting for it means that he has no sure method of assessing the mental capacity of extinct types of hominid simply by reference to cranial capacity.’
Since cranial capacity, per se, gives little indication of potential intelligence, evolutionists reason that the brain-body ratio would be a more accurate determinant of mental capacity.
Comparative brain-body ratios
The brain-body ratio (BBR), which is the ratio of brain weight to body weight, of a man is small (1:38–40), but other mammals have similar ratios. The bottle-nose dolphin has a 1:38 brain-body ratio, and a house mouse has a ratio of 1:40.10
Comparing humans with other primates, the supposedly closest relatives of man do not necessarily have the closest BBR to man. Starting with the chimpanzee, which has a BBR of 1:129,11 and a gorilla, which has a BBR of 1:200,12 the marmosets and squirrel monkeys, with BBRs of 1:19 and 1:12 respectively,12 show smaller ratios. Even though with such small ratios the latter two have large brains compared to their bodies no one would ever say that the latter two primates are more intelligent than the former two.
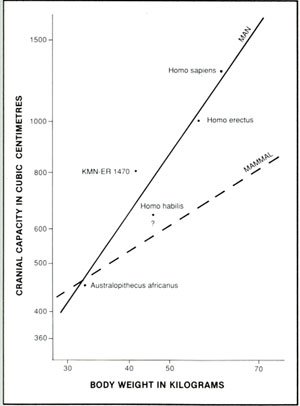
Figure 2. Brain-body ratio of fossil men and mammals.
Gould observed that the body weight and brain weight of mammals showed a consistent trend when plotted on a graph, called the ‘mammalian line’.13 He noted that between species if one triples the body weight of mammals the cranial weight doubles. One cannot acquire the cranial weight of fossil hominids, but since brain weight and cranial capacity are relatively the same, he could take fossil men, plot cranial capacity with body weight, and compare them to the mammalian line (see Figure 2). Gould states:
‘Some of our early ancestors, who were smaller than humans today, had brains closer to the average for their size of all mammals … this is Australopithecus africanus and later African Homo habilis … . The more recent, Homo erectus and modern Homo sapiens have much larger brains than predicted by body weight alone. This indicates that we have evolved larger brains at an exceptional rate and apparently have reached new levels of intelligence.’13
Thus, Gould recognized the distinct division between the Australopithecus and Homo habilis (recognized by most anthropologists as advanced A. africanus) and Homo erectus. The former two are on the mammalian line, whereas the latter is grouped with modern man. Thus according to BBRs creationists would divide the two groups into a general mammalian kind and a human kind.
Since it is difficult to show evolution by cranial capacity and brain-body parameters, more important recognition should be given to the direct observation of cranial configuration.
Comparative cranial anatomy
Essential to the study of endocranial configuration of fossil crania is the study of modern primate cranial anatomy. That determines what makes the human brain different from non-human primates? The brain of man shows a complexity that is far greater than the brain of any other of the non-human primates (see Figure 3).
The brain is divided into two hemispheres which are separated by a deep fissure, but bonded together by the corpus collosum. The cerebral cortex increases its area by gyri or furrows and sulci or fissures. The configuration of these fissures increases in complexity as one studies the primates from monkeys to man.
Roughly, the cortex is divided into frontal, parietal, temporal, and occipital lobes. The frontal and parietal are separated by the central sulcus, the parietal and occipital by the parieto-occipital sulcus, and the parietal and frontal from the temporal by the lateral fissure. According to Swindler and Wood,14 Homo is the only primate possessing a complete lateral fissure consisting of anterior horizontal, anterior ascending, and posterior rami (see Figure 3).
Homo shows an expansion in the frontal area, as evidenced by the central sulcus which lies posterior to the midpoint, whereas in gibbons and chimpanzees the sulcus is anterior to the midpoint.15
On the occipital pole, the lateral calcarine sulcus in monkeys and apes cuts across the occipital poles, but rarely in Homo. This sulcus is Y-shaped. Also, Barr16 says that in primate brains other than man (and in some human brains) there is a short calcarine sulcus which continues for a short distance over the occipital pole with a curved lunate sulcus known as the simian sulcus. A lunate sulcus is a trait that is either absent or ‘identified with difficulty’ in Homo.17 According to Figure 5, the Homo sulcus lunate when it exists in humans appears to be unique.
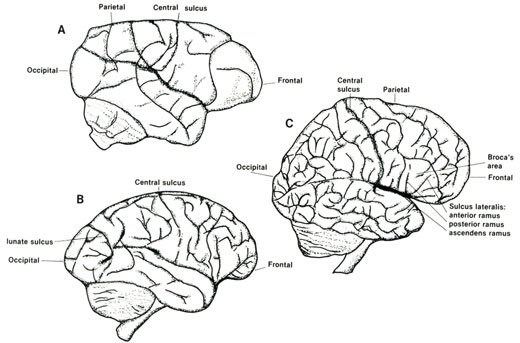
Figure 3. A comparative cranial
configuration of monkeys, chimpanzees, and man. A. Ceboid monkey B. Chimpanzee
and C. Man. Adapted from references 11 and 18.
The misuse of cranial configuration comparisons
In the early 1900s craniometricians (scientists who measured craniums to show the relationship between cranial capacity and intelligence) used cranial configuration and cranial area sizes to explain and support their theories. Paul Broca, who measured the cranial capacities of great and noble men to support his thesis that nobility had the highest capacity, was embarrassed by the measurement of the great mathematician K.F. Gauss who had an average brain of 1492cc. To explain the inconsistency, E.A. Spitzka showed that Gauss’s brain had many more convolutions compared to a Papuan’s brain which had fewer gyri.18
Broca and his colleagues also believed that higher mental functions were located in the anterior regions of the cortex, rather than in the posterior areas which dealt with processes such as involuntary movement, sensations, emotions, etc.18 He classified intelligent people as front thinkers and less intelligent people as back thinkers. Whites, he claimed, were frontal lobe thinkers, Mongolians middle thinkers and blacks back thinkers. Broca even had an explanation for the difference in configuration by stating that the differences were according to late or early suture closure.
Another person who explored differences of measurements of parts of the brain was Robert B. Bean in 1906. He felt that he had an intellectual ‘breakthrough’ with his statistics on the corpus callosum.18 Bean agreed with Broca’s ‘discovery’ that the front of the brain is used for higher reasoning and the back of the brain for sensorimotor activity. The corpus callosum has two parts: the genu in the front and the splenium in the back. He plotted the size of genus, claiming whites have a large genu and blacks had a larger splenium. He speculated that blacks would have a larger genu because they have a keener sense of smell since the genu contains the fibres of olfaction and intelligence. However, because Bean assumed that blacks had a smaller frontal lobe, they should logically have a smaller genu.18 In the early 1900s strong olfactory sense was associated with primitive man; thus Bean examined his data with a racial bias.
So Bean published in the American Journal of Anatomy in 1906, from which data he claimed that because of the constant relationship of the size of the brain and the size of the corpus callosum, there was support for the premise that whites are ‘frontal’ people and blacks are ‘occipital’ thinkers.18 He claimed that in the relative size of their frontal areas blacks are intermediate between the orangutan and true man.18 The problem with Bean’s assumptions is that he neglected to include sex in his statistics, since women, regardless of race, have smaller genus than men. Also, Bean said nothing about the total brain size of the individuals in his statistics and his data came from unclaimed bodies donated to medical schools.
Later, Franklin P. Mall at Johns Hopkins University repeated Bean’s studies with 106 brains and found no difference between whites and blacks in the size of their genus and spleniums. More important, then, are studies of the whole brain rather than just the studies of genus and spleniums.
The use of cranial configuration
Because of past abuse of cranial measurements and their relation to race, more importance is now being placed on endocranial configurations of fossil men. In earlier days cranial capacity was the most important measurement. For example, Marcellin Boule confronted Davidson and Black’s suggestions that Pekin Man was a primitive man because his 900cc of brain was not enough for any degree of intelligence. On the other hand, not all anthropologists feel that endocranial casts are an answer to the problem. Weidenreich so boldly warned:
‘… Therefore, the claims of paleoanthropologists for instance, to the effect that Neanderthal or Pekin man was right handed or left handed, was able to speak, or write or could only stammer, all deduced from shallow and narrow or deeper and broader impressions on the inside of the brain case, have no scientific basis even if the interpretation of the imprints could be accepted as correct’.19
Past studies of the evolution of mammals have not only shown an increase in the cranial capacity along the presumed pathway of evolutionary development, but also in some cases an increase in complexity. Tilly Edinger studied the supposed evolution of the horse and found that the neopallium showed more convolutions in the modern horse than Eohippus, but Weidenreich questioned if a complex brain really gave a more selective advantage for living. He argued that even though the North American horse had a well developed neopallium that was more complex than its progenitors, it still became extinct in the late Pleistocene. But then this horse did not exist on the earth any longer than its ancestors which had smaller, less complex brains, and in any case, Eohippus wasn’t a horse at all, its skeleton now being labelled as a hyrax.
Weidenreich went on to question the validity of the convolutions (wrinkles) when discussing the patterns on the surface of the hemisphere. On those areas, primates and man do not differ from other mammalian orders with regard to the presence and abundance of the wrinkle system. Some mammals have more complex wrinkles than primates. An example is the whale and its relatives which have the greatest wrinkles and the most intricate arrangements.
The capuchin monkey has almost a smooth brain, whereas the chimpanzee has a wrinkled one that comes closer to that of man. However, the chimpanzee’s convolutions, except for basic patterns, differ from the orangutans, gorillas and humans due to growth patterns. Weidenreich claimed that convolutions have an enormous variability not only in the length, breadth, and depth, but they also differ from person to person of the same race, even between right and left hemispheres in the same person.
Holloway concurred with this idea when he said that wrinkles and convolutions are not as important as the relative size of the frontal, temporal, parietal and occipital lobes as marked by the convolutions of the lobes themselves.20
Making endocranial casts of higher organisms has been a fruitless task in some instances. Symington in 1916 compared endocasts of modern human skulls with their wrinkles of the corresponding brains, and Clark in 193821 compared endocranial casts with chimpanzee brains. Both concurred that very little information about sulci patterns can be determined by endocasts. Holloway says that in virtually all cases the only detailed features that can be traced on the endocranial case of higher primates are the paths of the meningeal blood vessels.22 Indeed, the endocranial casts I’ve made (see Figure 4) also show that the meningeal arteries and the general shape of the brain are the only derivable information available.
Figure 4. A comparison of endocranial casts of a baboon, human and chimpanzee.
From left to right: Papio (baboon), Man, and Pan (chimpanzee).

Occipital view of casts.
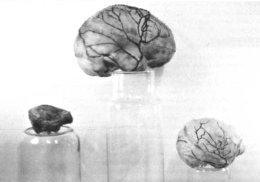
Right sagittal view. Note that the meningeal blood vessels are especially
prominent in humans. On the other hand, sulci patterns are difficult to see in human
endocranial casts.
Falk found, however, that although modern primates produce poor endocranial casts, many fossil hominids and other primates show sulci patterns.23 Furthermore, monkeys show convolutions on endocranial casts.
Some casts have been used to predict behaviour, such as Simons who says that the endocranial casts of Adapis show that it had a smaller brain size to body size than modern lemurs.24 Since Adapis had a larger olfactory bulb with less cerebral cortex in the occipital region, with larger temporal lobes, Simons concluded that Adapis had a greater allowance for smell than today’s lemurs. Another example is the endocast of Archaeolentur which showed transverse orientation of the sulci that resembled higher primates.
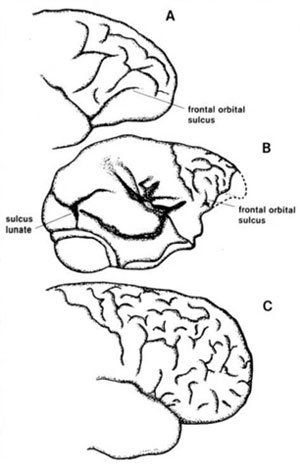
Figure 5. A comparison of the Taung child endocast
with apes and man. A. Frontal lobe of an ape. B. Taung specimen (A.
africanus). C. Frontal lobe of a human. The simplicity and what seems
to show sulcal pattern shows that the Taung resembles the chimpanzee.
Adapted from reference 5.
A prime example of how controversial endocranial casts can be involves the study of Australopithecus. The Taung child, Australopithecus africanus, was found in late 1924. Dart published a preliminary report of this find in Nature. In his report he stated that Taung was a human-like ape with features intermediate between living anthropoids and humans. He pointed out that the brain was large (525cc), but now estimated by endocranial cast as (405cc) and claimed that its general structure was more human than ape. In particular, the lunate sulcus, a groove on the rear portion of the brain which demarcates the visual portion of the brain, occupied a posterior position, as in humans (see Figure 5).
Holloway has always been in support of the interpretation that the Australopithecus had a brain close to humans. He says:
‘To recapitulate both the direct evidence of neurological organization and the indirect evidence of comparative brain size appear to indicate that Australopithecus and at least one other African primate of the same period (KNM-ER 1470) had brains that were essentially human in organization and that Australopithecus was also probably within the human range of sizes with respect to the proportion of the brain to the body.’25
But Falk disagrees with Holloway. He maintains that although human and chimpanzee casts tend to show meningeal arteries with very little definition, the casts of Australopithecus, especially the Taung, show details of the sulci on those organisms. Falk says that ape brains have a fronto-orbital sulcus that goes along the side of the frontal lobe to the lower surface. Humans have convolutions instead (see Figure 5). More important, Australopithecus lacks the sulci of the lower frontal lobe on the left side, known as Broca’s area, which controls speech. Falk claimed that in his endocranial analyses only KNM-ER 1470 and Homo reproduced a frontal lobe in what appears to be Broca’s area.26 Falk remarked at the end of his monograph:
‘The most important conclusion of this paper is that the australopithecine lunate sulcus was not located in a caudal human-like position, as first reported by Dart and now generally believed. Rather, the australopithecine lunate sulcus was relatively rostral, as in pongids.’27
Holloway responded to Falk’s reinterpretation of Australopithecus africanus by reiterating the claims of a hominid anterior placement of the lunate sulcus which Dart originally proposed. Today the debate still goes on between Falk and Holloway. Figures 6 and 7 compare the Taung child with an Indian child.

Figure 6. A comparison of the skull endocasts of a human, the Taung child, and adult chimpanzee. From left to right: Endocasts of a six-year-old American Indian child, the Taung specimen, and a chimpanzee. Top: The total Taung cast compared to a similar cast of the child along with an adult chimpanzee. Bottom: Endocasts of all organisms. The top and bottom chimpanzee specimens are of two different individuals. The flat of the back of the American Indian child is due to postnatal deformation. Note that the Taung specimen seems to resemble the chimpanzee more than the child.
Falk also has challenged another fossil man with his endocranial analyses. Proconsul africanus, found on Rusinga Island, Kenya, has had a history of controversy. Because of its sulcus markings on the frontal and parietal regions, Clark and Leakey claimed in 1951 that it was more cercopithecoid-like rather than hominoid.28 On the other hand, Radinsky in 1974 re-examined the casts and claimed the skull to be more gibbon-like rather than cercopithecoid, and therefore hominoid. Falk, upon examining the sulci, claimed that it had nine sulci which are basic anthropoid features on a small cranium, but a frontal sulcus and two other sulci are missing which are usually present in hominoids. Falk concluded that Proconsul appears to be a primitive anthropoid that has ceboid skull and dental characteristics but has mostly a gibbon cranial configuration. Proconsul is a ‘dental ape’, but neurologically he is a primitive anthropoid. So the first great apes’ configuration is in the australopithecine. So, although the cranial analysis of Proconsul has been in support of its being a transitional fossil ape to man, its cranial configuration studies have produced data in support of its affinities with other primates.
Conclusions
There have been many attempts to measure the cranial capacity of man and his cranial configuration in order to directly measure his mentality. From what is now known of modern man, there is no relationship between cranial capacity and intelligence. In fossil man and apes the endocranial casts show arteries and the general shape of the inner aspects of the skull, but not the sulci and gyri which are important. The key transitional fossils Proconsul and Australopithecus have been challenged by Falk and his group who demonstrate affinities of these organisms by ‘reading’ the sulci, especially the lunate sulcus on the endocranial casts. Their work is disputed and so one must conclude that cranial configuration studies need further research.
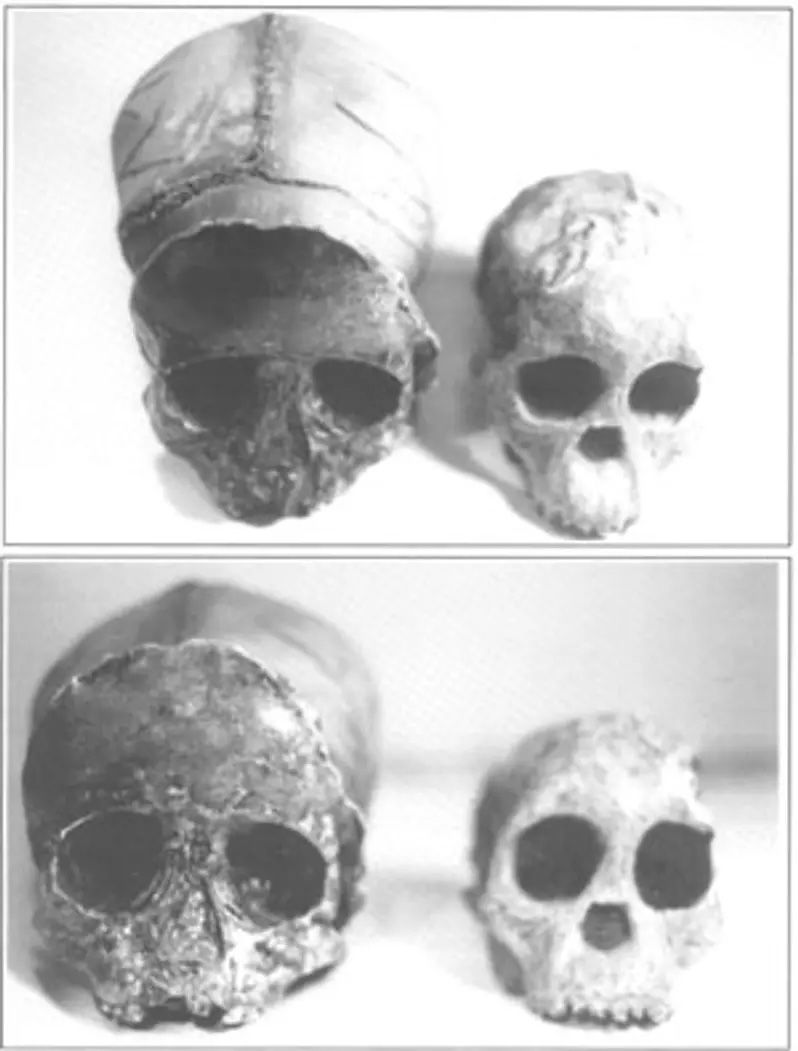
Figure 7. A comparison of the Taung endocast with an endocast of a human child of the same age.
Acknowledgment
I wish to acknowledge the contribution made by my daughter Bethany Jue in preparing the necessary line drawings for this article.
References
- Montague, M.F. Ashley, 1960. An Introduction in Physical Anthropology, Charles Thomas Press, Springfield, USA.
- Holloway, Ralph, 1974. The casts of fossil hominid brains, Scientific American 231(1):106ff.
- Olivier, George, 1969. Practical Anthropology, Charles Thomas Press, Springfield, USA, p. 135.
- Jerrison, Harry J., 1979. The evolution of diversity in brain size. In: Development and Evolution of Brain Size, Hahn, Jensen and Brududec (eds), Academic Press, New York.
- Schultz, A.H., 1966. The physical distinction of man. In: Readings in Anthropology, Thomas W. McKern (ed.), Prentice-Hall, Engle-wood Cliffs, New Jersey.
- Harris, Marvin P., 1971. Culture, Man and Nature. Thomas Y. Crowell, New York,
- Holloway, Ralph, 1966. Cranial capacity and neural reorganizations—a search for suitable parameters, American Anthropology 8:103.
- Weidenreich, Franz, 1967. The human brain in the light of the phylogenetic development in human evolution. In: Readings in Physical Anthropology, Noel Kom and Fred W. Thompson (eds), Holt, Rinehart and Winston Inc., New York.
- Clark, Wilfred H. LeGros, 1967. Bone of contention. In: Readings in: Physical Anthropology, Noel Kom and Fred W. Thompson (eds), Holt, Reinehart and Winston Inc., New York.
- Tobias, Phillip V., 1976. Brain size, gray matter and race—fact or fiction? In: Physical Anthropology and Archaeology. Peter B. Hammond (ed.), Macmillan Publishing Co., New York.
- Gould, Stephen Jay, 1974. Sizing up human intelligence. Natural History, 83(2): 14.
- Swindler, Daris R., and Wood, Charles D., 1973. Atlas of Primate Cross Anatomy: Baboon, Chimpanzee and Man, University of Washington Press, Seattle.
- Dillon, Lawrence, 1973. Evolution Concepts and Consequences, The C. V. Mosby Co., St Louis.
- Barr, Murray, 1974. The Human Nervous System: An Anatomical Viewpoint, Harper and Row, New York.
- Swindler and Wood, Ref. 12.
- Gould, Stephen Jay, 1981. The Mismeasure of Man, N. W. Norton and Co., New York.
- Gould, Ref. 16.
- Gould, Ref. 16.
- Weidenreich, Ref. 8.
- Holloway, Ref. 2.
- Falk, Dean, 1980. A reanalysisof the South African australopithecine natural endocasts. American Journal of Physical Anthropology, 53(4): 525.
- Holloway, Ref. 2.
- Falk, Ref. 21.
- Simons, Elwyn, 1972. Primate Evolution, Macmillan Publishing Co., New York.
- Holloway, Ref. 2.. p. 112.
- Falk, Dean, 1984. The petrified brain. Natural History, 93(9):36.
- Falk, Ref. 21, p. 538.
- Falk, Ref. 21.

Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis