

The Evolution of Feathers: A Major Problem for Darwinism
Originally published in Journal of Creation 17, no 1 (April 2003): 33-41.
Abstract
One of the most difficult issues related to bird evolution is the evolution of feathers. Feathers are complex, designed structures required for flight, and are today found only on birds.
Summary
The origin of birds has always been a major problem for Darwinism, and even today little agreement about the evolution of birds exists. One of the most difficult issues related to bird evolution is the evolution of feathers. Feathers are complex, designed structures required for flight, and are today found only on birds. A literature review on the evolution of bird feathers showed that even though feathers are found back as far as the Cretaceous, including many well-preserved samples in amber, the fossil record reveals a complete absence of evidence for feather evolution. The implications of this major difficulty for Darwinism are discussed.
Only birds and certain turkey-sized alleged theropods have feathers, no other animals do.1 Birds are ‘custom designed for flight’, one critical component of which is their feathers.2 The fact that birds use feathers to fly ‘affects virtually every aspect of their anatomy’.3 Feathers are the ‘most complex epidermal appendages found in animals’.4
These precision-designed structures are highly effective, yet extremely light, insulators that are designed to enable birds to fly. Bird wing feathers themselves are ‘one of the most beautifully designed structures in the world’.5 Feathers have such ‘striking diversity in size, shape, colours, and texture’ that very few human art masterpieces can compare with them in harmony of colours, liveliness and softness.4 Often copied but never equaled, the beauty in pattern and colours of feathers has been appreciated by humans the world over, as is evident from the fact that feathers have been used by humans to adorn themselves since ancient times.
The source of the colours
The black, brown and gray pigments in feathers come from the bird’s blood, and the red and yellow pigments from its fat. Lipochrome pigments produce red, orange and yellow colours, and melanin produces black, brown, red-brown and gray colours.6 The rainbow of colours on birds, including the blue shimmering on the throat and tail feathers, is due to both pigments and differential scattering of light. Referred to as structural colours, this method uses particles within the feather to scatter short wavelengths, because scattering intensity is proportional to the inverse fourth power of the wavelength. Also, microscopic ridges on the feathers act as a diffraction grating to break up the light that falls on them into all the colours of the light spectrum. These systems all combine to produce the vibrant rainbow of colours that birds commonly display. Their colours serve a variety of functions, including seemingly contradictory functions such as to attract mates and to blend in with their surroundings.
Many birds have thousands of feathers; a Plymouth Rock Hen has an estimated 8,000, and a Whistling Swan 25,000. Even a small bird such as a wren has over 1,000 feathers.7 Ogburn says:
‘Birds account for a disproportionate amount of our perception of nature not only because their flight, song and colours make them noticeable as well as appealing but because they are nearly ubiquitous.’8
The reason why they are so ubiquitous, Ogburn notes, is because, just as the airplane ‘has enabled men to extend their realm to the farthest reaches of the planet’, so the feather, the ‘masterpiece’ of nature, also has enabled birds to do so.8

The stunning array of colours is most apparent in the plumage of the peacock.
Types of integuments
All animals have some type of integument to maintain the integrity and integration of their bodies and protect them from pathogens, the weather and other adverse environmental conditions. The four basic types of integument structures that function to protect eukaryotes are:
Skin can be extremely thin, such as that covering many worms, of medium thickness, such as the skin covering humans, or thick and tough, such as the hide covering cows and elephants.
Many animals also possess some type of hair growth in or over their skin or hide. Most mammals are abundantly covered with thick hair (fur) for warmth and insulation.
Scales, such as those on reptiles, range from thin-but-strong scales covering snakes to thick iron-like scales protecting armadillos. Scales overlap like roof shingles and function in much the same way to keep water out and trap body heat while still allowing the animal to move.
Feathers are a unique structure found only in birds. All birds (class Aves), and no other animals, have feathers.9 Feathers are considerably different from scales and all types of skin, whether the skin is thin and hairy, or thick and hairless.
Feathers, scales, hair, fingernails and claws are all outgrowths of the skin, and are all as devoid of feeling as are animal hair or nails (for this reason, no pain is produced when hair is cut or nails are trimmed). Feathers and hair both form in pits called papillae located underneath the skin and often cover almost the entire body just as hair and scales do. Each pit is abundantly supplied with blood so as to nourish the growing feather. The feather, a strong-but-light structure, is constructed out of keratin, a strong-yet-flexible protein.
Most birds ‘shed’ or lose their feathers at regular intervals, usually once a year (a process called molting). Molting ‘is a highly orderly process’ that occurs gradually to insure no bare spots develop.6 It is so highly ordered that the ‘flight and tail feathers are lost in exact pairs, one from each side, so that balance is maintained’.6
The structure of a feather
The most common theory of feather origins suggests that feathers evolved from reptile scales. Before this topic is discussed, a review of feather structure is required. The anatomy of feathers is radically different from that of skin or scales, and is most similar to hair (yet much more complex). Bishop notes that: ‘Feathers may look simple, but they’re really very complicated. Each one can have more than a million tiny parts’.10 The complex anatomy of a feather varies, depending on the feather’s function. For example, the ‘complex morphology’ of the flight or contour feather includes a long shaft (usually hollow, always strong) and the web that flares out from the shaft in the form of roughly planar vanes on either side.11 The shaft consists of a hollow, stiff structure (often called a quill or vein) termed the rachis that serves as a solid-but-flexible support for branches called barbs. The rachis and barbs are analogous to a tree trunk and tree branches.12
Each parallel barb slants diagonally from the shaft, and has numerous smaller side branches or barbules (or webs) of different types that overlap those of the neighboring barbs in a herringbone pattern resembling a miniature replica of the whole feather.13 The flight feather of a large bird can have as many as a million barbules.14 The barbules in flying birds are held firmly to the next web by ‘hooklets’ called hamuli that function much like Velcro®. These branches and hamuli form a web sufficiently flexible, yet also stiff and dense enough so that when the bird flies, very little air or water is able to seep through the spaces.11 The branches and hooklets, in turn, contain barbicles.
The feather barbules also must be strong, yet flexible enough so that they will not break in the wind. Their design enables birds to ride air currents more gracefully than the best glider human engineers have ever designed. If the hooklets are lacking, such as in the plumes of the ostrich, the bird cannot fly. Hooklets are designed so that they can separate under certain conditions, preventing wind damage to the wing and feathers, but can be easily reattached when the bird preens its feathers (ruffled feathers normally prompt preening behavior). Humans can repair a ruffled feather simply by drawing it between their fingers. Preening behavior is part of the irreducible complexity of the total feather design.
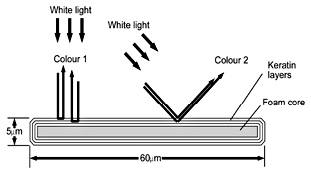
A cross-section of a peacock barbule showing the keratin layering which accounts for the iridescent colours (after Burgess).82
Feathers must be designed to give the bird ‘lift’ by causing the air on the top surface to flow faster then the air on the bottom surface, so that air pressure is lowered above the wing (the Bernoulli effect). To achieve this, flight feathers are asymmetric, with the smaller vane on the leading edge in direct contact with the air during flight. The ‘sophisticated aerodynamic principles in the design of the bird’s wing’ include a mechanism that reduces the adverse effect of turbulence—a major cause of airplane crashes.14 Specially designed slots in the bird’s airfoil cause part of the air stream to smooth out the airflow, an innovation imitated by aeroengineers in modern airplanes by designing small subsidiary airfoils in the wing. The bird also can vary its wing shape and flow traits to facilitate take off, flight control, and landing. One way of doing this is by an intricate system of tendons that allows the bird to twist its feathers so as to alter their resistance to air.14
Birds need feathers to fly, but feathers also give the bird much-needed protection against adverse weather, especially cold air and water. Overlapped like shingles and coated with a layer of oil, feathers protect the bird from water and heat loss much like roof shingles help to protect a house. As Peterson notes:
‘The feather is a marvel of natural engineering. It is at once extremely light and structurally strong, much more versatile than stretched skin on which a bat supports itself in flight, or the rigid structure of an aircraft’s wings—and far more readily repaired or replaced when damaged … . Though nearly weightless it has strength. The stiff shaft of the quill provides rigidity when support is needed, yet it is supple towards its tip, when flexibility is required for split second aerial maneuvering. Feel the sleekness of the web, soft yet firm. Separate the barbs; zipper them together again by running them through the fingertips as a bird would preen with its bill. The intricacy of the design that allows this can be appreciated by putting the feather under microscope.’15
Feathers are connected together, along with other structures, to form the plumage. The plumage, dermal and subdermal cutaneous muscles, ligaments, and the brain and sense organs form an ‘interconnected’ structure that must work as an irreducibly complex unit in order for the feather system to work at all. It is critical that even such details as the angle, thickness, shape and construction of all the feather parts are held to within narrow tolerances.16 Most minor deviations can render the entire system of flight unworkable. A theory of feather evolution also must account for ‘the structure and complexity of the follicle’.17
For this reason ‘it is not realistic to discuss the evolution of the avian feather independently of the evolution of the avian integument with all of its interconnected features’.18 Evolution of feathers (or any of the many other structures required to fly) as separate structures is unlikely and clearly counterproductive because, as separate structures, they would impede survival.
Special muscles on the skin enable birds to exercise detailed, controlled movement of their feathers. This system aids in flying, and sometimes in protection. Some birds fluff their feathers for various reasons, such as to give the appearance of more mass (to frighten enemies), to keep them warmer or to attract other birds during mating season. The famous feather spread of the male peacock is an excellent example of the high level of control that birds have over their feathers, as is their individual design.19 A further problem is that feathers are useless for flying (or most other functions) unless (and until) they are properly arranged on the bird. A clear pattern on the wing, tail and other parts of the bird is required.
The basic types of feathers
Over a dozen different kinds of feathers exist to achieve the many functions feathers serve, including not only flight but also signaling, courtship, waterproofing, streamlining the body, protective colourisation, insulation and even chemical defence.18 And none of these many kinds of feather variations leans in the direction of scales, as would be expected by Darwinists. Nor is there any evidence of transitional forms between any of the feather types. One of these variations is what is known as down feathers, where little or no shaft is present, and the barbs interlock far less often than in non-down feathers.
Powder-down feathers are down feathers that release talc-like powder to help waterproof the feathers and provide them with metallic-like luster.6Filoplume feathers contain hair-like projections on the end of each shaft that are used for decoration, courtship, sensory input and other purposes. In other types of feathers, such as the bristle feathers found on a flycatcher, the vein may be nearly or totally absent. Yet in other types, the vein may be solidified (such as on the penguin).
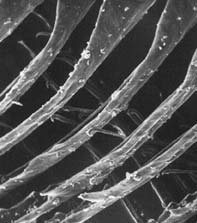
Courtesy David Menton.
An electron micrograph of a flight feather. The hook of the barbules can clearly be seen. Preening ‘resets’ these hooks into the proper position.
Yet another feather type is the flight or wing feathers (remiges), often called contour feathers, and tail feathers (rectrices) that function to help the bird guide its flight.9 Two basic types of flight feathers exist—fast and slow. The fast type are strong, trim feathers used in birds that travel at high speeds, such as pigeons and hawks. The slow type consists of a soft and loose-edged structure, and is used on birds such as owls that fly and soar at relatively slow speeds. Their advantage of slow feathers is that they produce flight that is far quieter than that allowed by fast feathers (quietness is far more important to an owl than speed).
The belief in feather evolution requires evidence for the evolution of each kind of feather (or evidence for the evolution of each feather from the first feather), which requires speculation about ‘feasible selective demands acting on evolution of feathers,’ a task no-one has yet achieved.18 The evolution of feathers is considered so improbable—even by evolutionists—that Darwinists generally conclude that ‘feathers evolved only once in the history of the vertebrata’.20 Much speculation also exists about this first feather—was it a simple contour feather, a downy feather or a flight feather?21
A major problem with feather evolution is ‘it is difficult to account for the initial evolution of feathers as elements in the flight apparatus, since it is hard to see how they could function until they reached the large size seen in Archaeopteryx’.22 In other words:
‘the chief difficulty in thinking about the evolution of the first feathers is the difficulty in accounting for the genesis of the structure through a continuous sequence of selective forces and with a continuous series of hypothetical morphological steps that are functionally plausible.’23
The common solution is to argue that feathers may have first evolved to provide insulation by a gradual increase in the size and the protofeather traits of scales.22 Known as the insulation theory, this idea has received a great deal of attention in recent years.
The insulation function of feathers
Since birds are endotherms (warm-blooded), to survive in cooler climates they must effectively stop much of their heat-loss—especially the loss from underneath their breast feathers. The air spaces between the feathers are highly effective insulators for the birds’ body. This function is so critical that many Darwinists have theorized that feathers actually originated primarily as an insulating device, and only later facilitated flight.9,24,25 The first step in bird evolution, therefore, is believed to be not the development of their ability to exploit the air, but of a light and effective ‘thermostat mechanism’ to maintain constant temperature—with part of this system later evolving into the feather.24
In contrast to this hypothesis, Parkes argues if ‘the primary “need” of the avian ancestor’ was for ‘an epidermal outgrowth … useful as a thermoregulatory mechanism … why “bother inventing” anything as complex as a feather? Hair would have been much simpler!’26 Indeed, a hair-like structure is better for insulation, which explains why flightless birds have feathers superficially similar to hair.27 In some cases this can be explained by loss of information for the complexity of flight feathers due to loss of selection pressure to maintain aerodynamic structure,25 so is compatible with the creation model.
Another problem is that ‘primitive’ down feathers are a poor means of temperature control (the flight feathers that use trapped air achieve much better insulation). Dewar concluded the insulating theory is erroneous because the assumption of a cold-blooded creature becoming warm-blooded is problematic for many reasons, including the fact that the transformation was supposed to have taken place in the tropics where temperatures are both fairly consistent and close to that of blood in warm-blooded animals (37ºC).28 For this reason, some argue that feathers evolved to help cool the bird by functioning as ‘sun shades’ that block the sun to reduce the level of heat absorption.29 Bock concludes that the thermal theory of feather evolution is at best only ‘a poorly tested theory’.30
Another problem that requires some explanation is the origin of the extremely complicated mechanism that birds use to avoid becoming overheated while in pursuit of their quarry. It is widely recognized that the heat-regulating mechanism of birds is poorly understood, and that no viable theory exists as to its origin. Problems include the fact that feathers reduce dissipation of the heat generated by the bird’s metabolism.
To overcome this problem birds have unique air sacs to assist in dissipating heat generated during periods of high activity. But these structures alone usually cannot maintain a constant temperature. Some speculate that, to help maintain a cool temperature, a complex nervous mechanism exists to regulate both the oxidation level and the amount of blood supplied to every organ. Evolutionary theories relating to the origin of feathers and flight (and even heat conservation) are all inadequate, and evidence for such an evolutionary origin is nonexistent.31
Since no evidence exists for any of these theories, they remain speculative at best, and in most cases are largely guesswork.
Many theories exist to explain flight evolution, including the gradual elongation of scales to produce a large surface for parachuting, then gliding, and finally flight (the tree-down or arboreal hypothesis). Tarsitano et al. show that from a functional, morphological standpoint, the tree-down theory is superior, and that serious problems exist with the major opposing model.16 Other authorities argue for the opposite hypothesis, called the ground-up or cursorial theory. Yale University professor John Ostrom, after showing why the tree-down theory is fatally flawed, argued eloquently for the ground-up theory, a position that he admitted was a distinctly minority view.32 But as often happens, yesterday’s heresy is today’s orthodoxy, and now the cogent arguments by the arboreal advocates against the cursorial theory are largely ignored.
Since no evidence exists for any of these theories, they remain speculative at best, and in most cases are largely guesswork (for a review of the history of bird evolution theories, see Ref. 3). As Carroll concludes, ‘neither structural nor physiological arguments have yet settled this controversy conclusively’,33 and many hypotheses are difficult or impossible to test. Regal concludes that all existing ‘theories relating the origin of feathers to flight … [are] inadequate’.31 Marden adds that ‘theorists have spent half a century fiercely debating whether avian flight evolved from ‘the trees down’, via gliding intermediates, or from ‘the ground up’, via running, leaping intermediates, with no resolution in sight’.34
Because of this problem, theories of feather origin have recently multiplied. Some researchers now argue that feathers evolved for water repellency, to function as a sink for excess sulfur waste, to function as a heat shield or to streamline the body for greater speed. But none of these theories come close to explaining their aerodynamic structure. Prum, in an extensive review of these theories, concluded that they are all ‘insufficient to explain the origin and diversification of feathers’, and added that they are actually a ‘hindrance to evaluating’ new fossil finds.35
The fossil record
The common assumption that birds do not preserve well because of their hollow bones is incorrect. They actually preserve very well in certain environments, especially lacustrine (lake) environments, inland water habitats and marine areas. As a result, bird fossils are common.36,37 A fairly abundant fossil record of both birds and feathers exists that enables us to draw some fairly firm conclusions about bird and feather history. Literally millions of impressions of a large variety of small animals—even insects and feathers—have been found in stone, clay, peat, tar and amber. The scales of dinosaurs and reptiles, the feathers of birds, the leaves of plants and even the wings of insects are all often clearly outlined in detail in the fossil record, enabling us to study these life forms in detail.
Feathers also are preserved by carbonized traces (present in about 70% of deposits), bacterial autolithification (conversion to rock involving bacteria), imprintation (such as Archaeopteryx), in coprolites (animal dung), and in amber, among other methods.37 In general, the better the fossil record in terms of preserving morphology for a class of life, the weaker the case for Darwinism for that class. In the case of animals for which only bone fragments are preserved, such as those fossils used to support human and whale evolution, the ambiguity of the bones from extinct forms has been wishfully interpreted as evidence for transitional forms.
Birds, as already mentioned, are actually ideal animals to use to study evolution because their fossils preserve very well. The fact that 9,000 living species are now known, all of which have a very unique skeletal morphology yet only 45 extinct bird taxa have ever been identified,36 providing strong evidence that relatively few types of non-modern birds have existed throughout history. This conclusion is supported by the fact that of 329 living families of terrestrial vertebrates, fully 79% have been found as fossils, as have 97.7% of the 43 living terrestrial vertebrate orders.38
The alleged fossil evidence for the evolution of feathers
What is found consistently in the fossil record is fully developed scales, feathers that are fully feathers, and skin that is clearly skin. No transitional structures consisting of feathers that are part feather and part scale, or even feathers that are less than modern types, have ever been uncovered.39 All of the earliest birds discovered, including Protoavis, have fully modern feathers: ‘the oldest known feathers … are already modern in form and microscopic detail’.40
Archaeopteryx, of which seven specimens or fragments have been confirmed so far, had perfectly developed ‘completely modern’ feathers that are ‘nearly identical with those of modern birds’, in spite of the fact that Archaeopteryx was a very different kind of bird compared to modern birds.41,42,43 Thus, Archaeopteryx ‘does not provide much information about the origin of feathers because its feathers are almost identical to those of living birds’.44 Furthermore, fully functional ‘feathers clearly existed prior to the existence of Archaeopteryx’.45 Archaeopteryx is important because until recently it was ‘the only direct evidence … of the earliest stages of avian evolution’.46 Other fossil feathers now date as far back as the Cretaceous.22
Extensive study of one Archaeopteryx feather, a 69-mm-long, perfectly preserved example that was dated at 150 million years old, is identical in all major details to modern feathers.47 As early as 1910, Pycraft concluded that the Archaeopteryx feather differed ‘in no way from the most perfectly developed feathers known to us’,48 and the wealth of discoveries since then has not altered this early view. Furthermore, we now have a large number of skin impressions from dinosaurs, and evaluations of the extant samples conclude that they are ‘unlikely to represent a predecessor to a feather bearing integument’.49
Attempts by Darwinists to hypothesize how feathers could have evolved have produced ‘a morass of contradictory theories and muddy thinking’.47 Early evolution textbooks, such as Osborn,50 discussed a set of transformations or ‘intermediate feather types’ that scientists were certain they would soon find in the fossil record. Yet so far, none has been found. Nonetheless, most evolutionists still theorize that feathers evolved from reptile scales.51 One early scientific account concluded that scales gradually became longer, fimbriated (bordered by filaments thicker than hairs), and ‘more and more efficient in the work of carrying the body through space’ until birds could fly with ease.52 The present scenario involves the elongation of reptilian scales, which then subdivide into a set of lateral plates that subdivide again to form the feather framework.53 These feather-evolution schemes, although they may appear plausible, all tend to obscure crucial difficulties, and are too vague to be able to criticize their specific claims.54


Courtesy David Menton.
The detailed structure of the feather (first picture) and scales (second picture) both magnified 80 times.
Since a major morphological difference exists between feathers and scales, a large number of functional transitional forms must have existed. Yet ‘little to nothing can be said from the fossil record on functions and roles of feathers, especially the intermediate stages between the reptilian scale and the primitive avian feather’.20 One major problem Klotz notes is the fact that feathers ‘cannot correspond to a whole scale but only to the outer half of the scale. The inner half or vascular core is believed to have atrophied’.55 Even speculating on the ‘most primitive stages of the evolution of feathers’ is very problematic.18
Feathers are not only ‘strikingly different from scales in their structure’, but their development path also is radically different.55 The many problems with scale-to-feather evolution have motivated the development of new theories of feather origins, such as their evolution from a ‘cylindrical epidermal invagination around the base of a dermal papilla’.41
Brush even concludes that feathers must have evolved from a conical shaped, tubercle-like follicle rather than a plate-like structure.56 One reason he argues for this view has to do with the fact that the most primitive feather must have a hair-like follicular mechanism to produce feather proteins, which then must be properly assembled to produce the many molecular structures needed to form the feather’s complex gross anatomy. Therefore, the complex follicular mechanism must have evolved first. Furthermore, feathers have many more similarities, both morphologically and biochemically, to hair than scales.57
Although no evidence exists for this primitive follicular structure in the abundant bird fossil record, Brush’s theory does demonstrate a major failure of current scale-to-feather evolution theories. Hair-to-feather evolution is actually the most logical theory for many reasons, including the fact that a hair follicle already exists. The major argument against this theory is that birds are speculated to have evolved from reptiles, not from mammals. Therefore, most all researchers have totally ignored the hair-to-feather theory.
Feathers in amber
One of the oldest feathers, found in amber, dates back to the Cretaceous and is ‘an almost complete beautifully preserved’ feather. This example is a fully developed semiplume that is located at the margin of feather tracts and the apteria (the naked spaces between the feathered areas of birds) and usually is overlain by the contour feathers. The rachis and barbs were perfectly formed, and it sometimes is possible to identify the bird from which the amber-entombed feather has come. Unfortunately, many specimens have not yet been carefully studied. One that was identified was from a Pididae (woodpecker). Further studies no doubt will aid in the identification of many others. Although many fossil feathers and amber-preserved feathers (some dating back to the Cretaceous) have been located, no clues of feather evolution have ever been found in the fossil record—and consequently no physical evidence exists of the many changes that feathers must have undergone if they evolved.
In the words of one Columbia University biologist, ‘we lack completely fossils of all intermediate stages between reptilian scales and the most primitive feather’.20 So far, not one of the required, hypothetical transitional types has been discovered in the abundant bird fossil record. Millions of fossil birds exist in the fossil record, and all of them have perfectly formed feathers. For this reason, feather origin is considered an enigma by Darwinists.58
A major problem in feather evolution
A major problem with all Darwinistic theories of both feather and bird evolution is the fact that functional integrity is required for life, just as it is for any complex machine, involving not only the feather but also its many support structures (including the follicle, muscle, and nervous systems). In other words
‘ … organisms at every stage in the evolutionary sequence must be functional wholes interacting successfully with selective demands arising from the particular environment of the organisms at each stage in the evolutionary sequence.’59
Flight requires not only the evolution of feathers, but also a total redesign of almost the entire animal. In chicken embryos, feather development begins at day 6 and is intimately connected to the bird’s whole development.60 As a result, birds are ‘the most clearly distinct of all vertebrate classes’, and there is an ‘enormous gap in anatomy, and way of life’ between birds and their putative closest relatives, the reptiles.61
For example, the ‘metabolic rate and sustained body temperature are higher than in all other’ vertebrates. Carroll concludes that the ‘geometry and mechanics of their respiratory system are unparalleled’.61 Indeed, while reptiles have bellows-like lungs, birds have a complicated system of air sacs which keep air flowing in one direction through special tubes (parabronchi) in the lung, and blood moves through the lung’s blood vessels in the opposite direction for efficient oxygen uptake,62 an excellent engineering design.63 A related problem is that ‘the sequence of evolutionary steps must be continuous and gradual with no large saltational changes’.59 These two problems have proved lethal to all past and present theories of both feather and flight evolution. As an evolutionary respiratory physiology expert at Oregon State University in Corvallis, argues:
‘Recently, conventional wisdom has held that birds are direct descendants of theropod dinosaurs. However, the apparently steadfast maintenance of hepatic-piston diaphragmatic lung ventilation in theropods throughout the Mesozoic poses a fundamental problem for such a relationship. The earliest stages in the derivation of the avian abdominal airsac system from a diaphragmatic-ventilating ancestor would have necessitated selection for a diaphragmatic hernia [i.e. hole] in taxa transitional between theropods and birds. Such a debilitating condition would have immediately compromised the entire pulmonary ventilatory apparatus and seems unlikely to have been of any selective advantage.’64
This was in the context of the fashionable theropod origin of birds, but it seems to an equally effective criticism of all reptile-to-bird origin theories.
New discoveries
It is still much too early to evaluate the numerous recent claims of feathers on dinosaurs or other animals, some of which have been shown to be forgeries. Others also may prove to be forgeries, while some may offer very debatable evidence for feather evolution.65 In China (the source of all of the new feather finds), one researcher claimed ‘assembly line factories’ exist to assemble fossil forgeries that sell for large sums of money.66 The most infamous of these is Archaeoraptor liaoningensis, which was proven by computerized tomography scans to be a composite of several fossils.67,68,69
Many of the alleged ‘intermediates’ actually are fully formed, modern feathers, or structures that are not feathers at all.65 For example, Sinosauropteryx ‘feathers’ are actually ‘filaments’.70,71
Other ancient fossil feather discoveries—such as on the oviraptorosaur Caudipteryx and Protarchaeopteryx—are ‘true feathers’.72 Touted by some as a dinosaur, Caudipteryx has been dated to within the early Cretaceous, often estimated as 30 million years younger than Archaeopteryx.73 Evolutionary paleo-ornithologists Feduccia and Martin, staunch critics of the dinosaur-to-bird theory, believe that Protarchaeopteryx and Caudipteryx are more likely to be flightless birds similar to ostriches. They have birdlike teeth and lack the long tail seen in theropods. Caudipteryx even used gizzard stones like modern plant-eating birds, but unlike theropods. Far from being ancestors of Archaeopteryx, cladistic evidence points (under evolutionary presuppositions) to their being birdlike (under their own transforming paradigm), and secondarily flightless descendants of Archaeopteryx.74
Other new discoveries—such as the hair like filamentous integumental appendages on Sinornithosaurus millenii, a non-avian dinosaur—have only complicated the Darwinian theory (such as discussed by Zhang and Zhou44).
Conclusions
Even though fossil impressions of feathers are abundant in the fossil record, and much has been written speculating on how scale-to-feather evolution could have occurred, not a shred of fossil or other evidence has ever been found to support the scale-to-feather evolution theory.1,23 In the words of Prum, understanding ‘the evolutionary origin of feathers has been constrained by the lack of any known ancestral feather morphologies or structural antecedents’.41
The evidence supports Klotz’s early conclusion that the ‘origin of feathers is still a real problem’ for Darwinism, and all contemporary theories of feather origin are hypothetical ideas that ‘can only be characterized as judicious speculation’.75 In short, nothing has changed since Regal stated ‘although most textbooks include some sort of speculation on the evolutionary origin of feathers … [a] morass of contradictory theories and muddy thinking … occurs in … much of the literature on this subject’.31
Although much speculation and major disagreements exist on how feathers ‘could have’ evolved, all existing theories are ‘just-so stories’, unsupported by fossil or historical evidence. The profound evolutionary enigma of feathers noted by Darwin76 and Heilmann77 remains, even today. The lack of evidence for feather evolution is not only a major problem for Darwinism, but the design and function of feathers provides evidence for both intelligent design and irreducible complexity. Flight and feathers are indeed a ‘miracle’.78 Feather evolution is related to the question of bird evolution.
Periodically, new bird fossils are found, but most of them have been of little or no use as evidence of bird evolution, and the few claimed examples typically generate much debate. For instance, Feduccia concluded that one recent find, known as Apsaravis, contributes little
‘… to our understanding of avian evolution, and its lack of a clear relationship with any kind of modern bird makes its significance ambiguous. If Apsaravis is not related to any modern ornithurine, how can it tell us anything important about the evolutionary questions raised by [its discoverers] Norell and Clarke?’79
The latest discovery of feathers on the birdlike, turkey-sized ‘theropods’ Caudipteryx and Protarchaeopteryx indicate that they are flightless birds. Much debate exists about this and related finds.80 Some consider these animals to be birdlike dinosaurs, or other dinosaur-like, flightless birds that have lost their full flight plumage (or never developed it). Conclusions on these finds will require much more study, and yet already have produced much debate and controversy.
Much disagreement still exists about Archaeopteryx, a discovery now around 150 years old. Likewise, the place in evolution, if any, of the recent finds may never be settled. Many of these finds are from a province of China, and already one find from this area has proven to be a hoax. Consequently, much more study is necessary to determine the value of these finds. So far, none of these finds challenges the conclusions presented in this paper, and early study of these finds has strongly supported the findings reviewed here.
In conclusion, we agree with Brush: ‘Uncountable numbers of words have been written in attempts to … reconstruct the primitive feather and explain why feathers evolved’.56 So far, all of these attempts have not only failed, but also have led to the conclusion that how feathers ‘arose initially, presumably from reptilian scales, defies analysis’.81
Acknowledgments
I wish to thank Clifford Lillo and Bert Thompson for reviewing an earlier draft of this paper.
Footnotes
- Parkes, K., Speculations on the origin of feathers, Living Bird 5:77–86, 1966; p. 77.
- Bishop, N., The Secrets of Animal Flight, Houghton Mifflin, Boston, p. 8, 1997.
- Witmer, L., The Search for the Origin of Birds, Franklin Watts, New York, p. 9, 1995.
- Prum, R.O. and Williamson, S., Theory of the growth and evolution of feather shape, J. Experimental Zoology (Molecular, Developmental, Evolution) 291:30–57, 2001.
- Compton, Comptons Encyclopedia, The University of Chicago, Chicago, Vol.4, 5:56, 1957.
- Hickman, C., Roberts, L. and Larson, A., Integrated Principles of Zoology, McGraw-Hill, New York, p. 588, 2001.
- Taylor, B., The Bird Atlas, Dorling Kindersley, New York, p. 5, 1993.
- Ogburn, C., The Adventure of Birds, William Morrow, New York, pp. 66–67, 1957.
- Bock, W.J., Explanatory history of the origin of feathers, American Zoology 40:478–485, 2000.
- Bishop, Ref. 2, p. 9.
- Bock, Ref. 9, p. 478.
- Terres, H., The Audubon Society Encyclopedia of North American Birds, Alfred A. Knopf, New York, 1980.
- Peterson, R.T., The Birds, Time, New York, 1963.
- Denton, M., Evolution: A Theory in Crisis, Adler and Adler, Bethesda, p. 202, 1986.
- Peterson, Ref. 13, p. 33.
- Tarsitano, S.F., Russell, A.P., Horne, F., Plummer C. and Millerchip, K., On the evolution of feathers from an aerodynamic and constructional view point, American Zoology 40:676–686, 2000.
- Prum, R.O., Development and evolutionary origin of feathers, J. Experimental Zoology (Molecular, Developmental, Evolution) 285:291–306; 292, 1999.
- Bock, Ref. 9, p. 479.
- Burgess, S., The Beauty of the Peacock Tail and the Problems with the Theory of Sexual Selection, TJ 15(2):94–102, 2001.
- Bock, Ref. 9, p. 480.
- Bock, Ref. 9, p. 483.
- Carroll, R., Patterns and Processes of Vertebrate Evolution, Cambridge University Press, New York, p. 314, 1997.
- Regal, P., The evolutionary origin of feathers, The Quarterly Review of Biology 50(1):35–66, 1975; pp. 35–36.
- Tucker, B.W., Functional evolutionary morphology: the evolution of bird; in: DeBear, G.R. (Ed.), Evolution, Clarendon Press, Oxford, pp. 330–332, 1938.
- Ostrom, J.H., Archaeopteryx and the origin of flight, Quarterly Review of Biology 49(1):27–47, 1974.
- Parkes, Ref. 1, p. 81.
- Feduccia, A., The Origin and Evolution of Birds, 2nd Ed., Yale University Press, New Haven, p.130, 1999.
- Dewar, D., The Transformist Illusion, DeHoff, Murfeesboro, 1957.
- Bock, Ref. 9, p. 481.
- Bock, Ref. 9, p. 484.
- Regal, Ref. 23, p. 35.
- Ostrom, J.H., Bird flight: how did it begin? American Scientist 67:45–56, 1979.
- Carroll, Ref. 22, p. 314.
- Marden, J., How insects learned to fly, The Sciences 35(6):26–30, 1975; p. 27.
- Prum, Ref. 17, p. 292.
- Davis, P. and Dyke, G., Birds; in: The Encyclopedia of Paleontology, Fitzroy Dearborn, Chicago, p. 162, 1999.
- Davis, P. and Briggs, D., The fossilization of feathers, Geology 23(9):783–786, 1995.
- Denton, Ref. 14, p. 189; data compiled from Romer, A.S., Vertebrate Paleontology, 3rd Ed., University of Chicago Press, Chicago, pp. 347–396, 1966.
- Stahl, B.J., Vertebrate History: Problems in Evolution, Dover, New York, p. 350, 1985.
- Martin, L. and Czerkas, S.A., The fossil record of feather evolution in the Mesozoic, American Zoology 40:687–694, 2000; p. 687.
- Prum, Ref. 17, p. 291.
- Carroll, Ref. 22, p. 315.
- Cousins, F.W., The alleged evolution of birds; in: Patten, D. (Ed.), A Symposium on Creation, Vol. III, Baker, Grand Rapids, Chapter 4, 1971.
- Zhang, F. and Zhou, Z., A primitive enantiornithine bird and the origin of feathers, Science 290:1955–1959, 2000; p. 1957.
- Brush, A.H., Evolving a protofeather and feather diversity, American Zoology 40:631–639, 2000; p. 632.
- Ostrom, Ref. 25, p. 27.
- Parkes, Ref. 1, p. 77.
- Pycraft, W.P., Animal Life: An Evolutionary Natural History, Vol. II—A History of Birds, Methuen, London, p.39, 1910.
- Martin and Czerkas, Ref. 40, p. 687.
- Osborn, H.F., The Origin and Evolution of Life, Charles Scribners, New York, p. 228, 1918.
- Maderson, P.F., On how an archosaurian scale might have given rise to an avian feather, American Naturalists 146:424–428, 1972.
- Pycraft, Ref. 48, p. 39.
- Maderson, P.F. and Alibardi, L., The development of the sauropsid integument: a contribution to the problem of the origin and evolution of feathers, American Zoology 40:513–529, 2000; p. 514.
- Denton, Ref. 14, p. 216.
- Klotz, J., Genes, Genesis and Evolution, Concordia, St. Louis, p. 460, 1970.
- Brush, Ref. 45, p. 631.
- Sawyer, R.H., Glenn, T., French, J.O., Mays, B., Shames, R.B., Barnes, Jr., G.L., Rhodes, W. and Ishikawa, Y., The expression of beta (ß) keratins in the epidermal appendages of reptiles and birds, American Zoology 40:530–539, 2000.
- Turner, C.E.A., Archaeopteryx, a bird: no link, Evolution Protest Movement, Sept. 1973.
- Bock, Ref. 9, p. 482.
- Chuong, C.-M., Patel, N., Lin, J., Jung, H.-S. and Widelitz, R.B., Sonic hedgehog signaling pathway in vertebrate epithelial appendage morphogenesis: perspectives in development and evolution, Cellular and Molecular Life Sciences 57:1672–1681, 2000.
- Carroll, Ref. 22, p. 306.
- Schmidt-Nielsen, K, How birds breathe, Scientific American, pp. 72–79, December 1971.
- Engineers make much use of this principle of counter-current exchange which is common in living organisms as well—see P.F. Scholander, The Wonderful Net, Scientific American, pp. 96–107, April 1957.
- Ruben, J.A., et al., Lung structure and ventilation in theropod dinosaurs and early birds, Science 278(5341):1267–1270, 1997.
- Dalton, R., Feathers fly over Chinese fossil birds legality and authenticity, Nature 403:689–690, 2000.
- Dalton, R., Chasing the dragons, Nature 406:930–932, 2000; p. 932.
- Sloan, C., Feathers for T. rex? National Geographic, pp. 98–107, Nov. 1999.
- Dalton, Ref. 66, pp. 689–690.
- Editorial, Disappearing discovery of the year: Archaeoraptor, Science 290:221, 2000.
- Brush, Ref. 45, p. 632.
- Rossmann, T., Feathered or furry dinosaurs, TJ 16(3):19, 2002.
- Padian, K., Dinosaurs and birds—an update, National Center for Science Education Reports 20(5):28–30, 2000; p. 30.
- Swisher, C.C., III, Wang, Y-q., Wang, X-l., Xu, X. and Wang, Y., Cretaceous age for the feathered dinosaurs of Liaoning, China, Nature 400:58–61, 1999.
- Woodmorappe, J., Bird Evolution: Discontinuities and Reversals, TJ 17(1):88–94, 2003.
- Klotz, Ref. 55, p. 460.
- Darwin, C., The Origin of Species, John Murray, London, 1859.
- Heilmann, G., The Evolution of Birds, Witherby, London, 1926.
- Cromer, R., The Miracle of Flight, Doubleday & Company, Garden City, New York, 1968.
- Feduccia, A., Fossils and avian evolution, Nature 414:507–508, 2001; (with Mark A. Norell and Julia A. Clark replying on p. 508).
- Xu, X., Zhou, H-h. and Prum, R.O., Branched integumental structures in Sinornithosaurus and the origin of feathers, Nature 410:200–204, 2001.
- Stahl, Ref. 39, p. 349.
- Burgess, S., The Beauty of the Peacock Tail and the Problems with the Theory of Sexual Selection, TJ 15(2):94–102, 2001.

Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis