
Chapter 3
Beneficial Aspects of Bacteria
Section 3
by
Dr. Alan L. Gillen
on
August 14, 2015
Republished with permission and featured in
The Genesis of Germs
As Pasteur first pointed out, it would be difficult to overemphasize the importance of the activities of saprophytic microbes in decomposing the bodies of dead animals, plants, and non-living organic matter of all sorts. The final result of all the activity of this “rotting” bacteria is the conversion of entire complex organic material into simple, inorganic substances. The process of decomposition itself is not pleasant to contemplate. A piece of putrefying meat or a rotten egg, with its disagreeable appearance and foul odor, is a disgusting object, but it takes little imagination to realize the vital importance of the rotting process. Suppose the dead bodies of plants and animals, and fecal matter and other waste materials excreted from man and animals, accumulated where they fell and never decomposed! There would soon be no room on the earth’s surface for living things. As Pasteur said, the earth would soon be “encumbered with cadavers.” In decomposing lifeless, organic remains, bacteria make the earth fit for the living. Pasteur was able to comprehend the Creator’s wise plan for recycling the “dust of the earth.”
The decomposition of non-living materials by bacteria not only rids the earth of useless waste but also accomplishes an even more important result. It releases from dead or lifeless matter the elements needed for the growth of plants, returning these essential elements to the soil. The activities of saprophytic bacteria furnish the link between the dead and the living. All-important nitrogen is locked up in the bodies of dead plants and animals in the form of organic compounds. Nitrogen is released from its complex combinations by the action of putrefactive bacteria, passing into the soil in the form of nitrates—simple, inorganic compounds that plants use for food. Thus, nitrogen, really the master element of living substances, passes through a perfect cycle from the soil into the growing plant, then to the animal body when the plant is eaten, and back again to the soil through the agency of bacteria when the plant or animal dies. Other elements necessary for life, such as carbon, phosphorus, and sulfur, pass through a similar cycle in which bacteria play an important part. The supply of these chemical elements on earth is limited, and soon would be exhausted if not for the necessary work of bacteria. The fertility of the soil is, therefore, largely a result of bacterial activity.
The Nitrogen Cycle
Certain kinds of bacteria have an especially marked influence on soil fertility because of their participation in the nitrogen cycle (below). These are the nitrifying, denitrifying, and nitrogen-fixing bacteria. Nitrifying bacteria bring about the oxidation of ammonia (NH3) to nitrites (-NO2) and of nitrites to nitrates (-NO3). This is called nitrification. Since nitrates are the most useful form of nitrogen for the nutrition of plants, the amount of nitrates in the soil largely determines its fertility. The action of the nitrifying bacteria thus directly enriches the soil. These organisms are strictly autotrophic and non-spore-forming. Some are bacilli; others are cocci or spirilla. Nitrosomonas oxidizes ammonia to nitrites, and changing nitrites into nitrates is accomplished by Nitrobacter. A few other types of bacteria and fungi can also bring about the oxidation of ammonia or nitrites.
Denitrifying Bacteria
The action of the denitrifying bacteria is to bring about the reduction of nitrates (-NO3) to nitrites (-NO2), and the release of free ammonia (NH3) and sometimes free gaseous nitrogen, as the final steps in the microbial decomposition of lifeless organic matter. Various anaerobic soil organisms utilize nitrates as hydrogen acceptors, thereby reduced from nitrates to nitrites. Proteins and other organic nitrogenous compounds are decomposed to amino acids, and the amino (-NH2) groups are split off in the form of ammonia (NH3). Urea, a metabolic waste product found in the urine of man and animals, is decomposed by various microorganisms, with liberation of ammonia. These elementary, reduced nitrogen compounds—nitrites and ammonia—are useless to growing plants. They would be largely lost to the nitrogen cycle if not for the oxidizing capacity of the nitrifying bacteria.
Nitrogen-Fixing Bacteria
A most important contribution to soil fertility is made by still another group of bacteria—the nitrogen-fixing bacteria. These organisms can fix nitrogen from the air; i.e., capture atmospheric nitrogen gas and cause it to combine with other chemical elements to form organic compounds in the soil, eventually becoming available in a form useful for the nourishment of plants. Thus, free nitrogen in the atmosphere is put to work. This remarkable nitrogen-fixing property is possessed by two groups of soil bacteria.
The members of one group live free in the soil. An example is the organism named Azotobacter. These free-living organisms utilize carbohydrates in the soil as a source of energy and obtain nitrogen directly from the air. The nitrogen is combined into their protoplasm and later released in the form of nitrates, or other usable compounds, when the organisms die. Bacteria are themselves decomposed by other soil microbes. The growth of Azotobacter also enriches the soil with another needed element—phosphorus—which, like nitrogen, is built into the growing cells. Bacteria of some other genera (e.g., Clostridium and Aerobacter) also can fix atmospheric nitrogen.
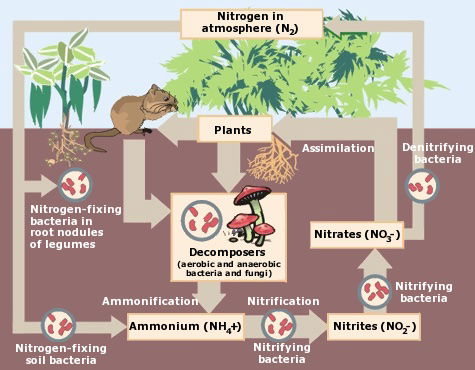
The Nitrogen Cycle (image credit, EPA)
The second group of nitrogen-fixing organisms is composed of root-nodule bacteria, which live symbiotically in nodules on the roots of certain plants (right). The plants that bear the nodules on their roots are called legumes. They include clover, peas, beans, and alfalfa. It has been known for centuries that plants of this kind enrich the soil. Farmers have long rotated crops on a particular field, sowing clover, soybeans, or some other leguminous plant every so often. When the clover is plowed under and the field planted again, the land is found to be more fertile. It is a common practice to inoculate soil or seeds with these organisms before planting to be certain that active development of the root-nodule, nitrogen-fixing bacteria occurs.
Rhizobium leguminosarum is a principal species of root-nodule bacteria. The rhizobia enter the tips of the root hairs of leguminous plants. They become surrounded by a kind of gelatinous coating or capsule and penetrate further into the roots. The organisms extend their growth into the tissue cells of the plant, and these cells respond by proliferating to form a protective nodule. The organisms in these root nodules utilize carbohydrates and other nutrients from the plant juices, simultaneously fixing nitrogen directly from the air. The end result of this remarkable symbiosis is the addition of large amounts of nitrogen to the soil.
Creation Scientist Focus 3.3
Microbiologist Joseph Francis
The Biomatrix of Life: A Creationist Perspective of Microbes and Viruses

Dr. Joseph W. Francis is a cellular immunologist and currently an associate professor of biology at The Master’s College in Santa Clarita, California. He taught biology at Cedarville University in Ohio for ten years prior to his position at Master’s. While at Cedarville, he was named faculty scholar of the year in 2000. He has a B.S. degree in microbiology from Michigan State University and a Ph.D. in biology from Wayne State University. After graduate school, he accepted an appointment as a post-doctoral fellow at the University of Michigan in the department of pediatric hematology and worked in cellular immunology. Dr. Francis has published over 20 scientific papers and given numerous presentations at scientific meetings. He has also authored and teaches an online, college-level biology course. He is a member of the Society for Leukocyte Biology, as well as the Baraminology Study Group, a society of creation biologists dedicated to developing a creation model built upon the truth claims of the Bible.
In his baraminology studies, Dr. Francis is developing models attempting to explain and describe how microbes and viruses may have functioned in the perfect pre-Fall environment as described in Genesis. Dr. Francis has discovered that a remnant of the original “very good” creation can be observed in microbes and viruses today. For instance, only a small fraction of microbes and viruses cause pathology and disease. In fact, a large percentage of microbes appear to beneficially interact with and positively affect biological life in numerous ways. For instance, microbes are involved in all major biogeochemical cycles, including the carbon and nitrogen cycle. Dr. Francis has referred to this pervasive functional population of the microbes on earth as a “biomatrix” or “organosubstrate.”
Dr. Francis proposes that microbes and viruses were created as a link between macroorganisms and a chemically rich but inert physical environment, providing a substrate upon which multicellular creatures can thrive and persist in intricately designed ecosystems. Consistent with this perspective, microbes and viruses are abundant in all ecosystems, separated by the discontinuity from macroorganisms that separates the major groups of viruses and microbes, designed for symbiotic relationships with both macroorganisms and other microbes and viruses, extract inorganic minerals from earth minerals, participate in the cycling of all elements and compounds important in macroorganismal biology, and effect bioremediation. This concept explains the organelle/bacterial similarities used as evidence of evolutionary endosymbiosis theory. The organosubstrate concept also suggests that pathogenesis is a relatively recent and rare deviation from original created function. Evidence of this includes the rarity and lower fitness of pathological forms, and the late addition to and modification of symbiotic design features in microbes and viruses. The organosubstrate concept is imminently testable and well-supported.
Furthermore, all living creatures harbor microbial symbionts. These microbial symbionts are primarily involved in providing nutrition for organisms, and also may be involved in reproduction control and host defense reactions. It also appears that microbes were created to interact with biological organisms, including humans. Dr. Francis is working on a theory that would show that microbes and viruses possess design features consistent with extracellular or extraorganismal organelles. We certainly have no problem accepting the fact that our cells have intracellular organelles, therefore we shouldn’t be surprised about the existence of extracellular organelles. There are many potential advantages to possessing such organelles. For instance, they perform many functions that would be burdensome or energetically costly to their hosts. Because of their independent nature, they could leave the host but be picked up from the surrounding environment, possibly providing information promoting the host’s adaptation to it.
This new way of looking at microbes and viruses also raises many questions about the immune system. If there was no death, disease, and putrefaction in the garden, what was the original role of the immune system? We certainly cannot rule out the idea that God knew about the fallen environment in which we would live, and therefore in His foresight and mercy created this system. However, it appears the immune system possesses specific, complex mechanisms that recognize and adhere to microbes and viruses. They are so complex that we can hypothesize that the immune system could have been created for more than just host defense. Perhaps the immune system was created to aid in the uptake and harboring of microbial and viral symbionts that God intended to be part of human bodies, animals, and plants. Modern medicine has recently shown that we experience many health problems when the normal flora bacteria in our intestines are compromised. In fact, the intestinal tract does not develop correctly in animals in which bacteria have been removed. Recent studies have shown that the immune system interacts with normal intestinal flora, and may control its population and promote its maintenance there. In this association between the immune system and normal flora bacteria in the intestine, we may be witnessing a snapshot of the original purpose of the immune system.
Cyanobacteria
The Cyanobacteria (blue-green algae) are microscopic—the simplest and most bacteria-like of the algae. Cyanobacteria are incredibly tolerant to harsh environmental conditions, thriving in waters high in nutrients or fertilizer runoff. Some can fix atmospheric nitrogen from the environment in cells called heterocysts. One example is Anabaena (right), which contains enzymes that fix nitrogen (N2) into ammonium (NH4) for use by the growing cell. Species growing in water usually have gas vacuoles that help the cell float in favorable environments. It is therefore not surprising to find among them forms adapted to growth in almost every sort of environment, including places in which no other kind of vegetation can exist. They occur not only in fresh or brackish water at ordinary temperatures, but also in hot springs at temperatures of up to 80°C (176ºF), in cold mountain streams, and even in water with a salt concentration as high as 27 percent. The bright colors of the terraces and “paint pots” around the hot springs at Yellowstone are due to the blue-green algae in the outer layers of this calcareous deposit. Other varieties may be visible to the naked eye as a moss-like film on virgin or cultivated soil, and still others are found as parasites, or in symbiotic association with various higher plants and animals.
The blue-green algae may grow as individual microscopic cells somewhat larger than bacteria, but they often form long, filamentous colonies consisting principally of multiples of just one kind of algal cell held together within a gelatinous sheath. When a heavy spreading growth of these algae develops, as, for example, over the surface of a pond or reservoir, or over a sizable area in the sea, this is called a bloom. There are no organs of locomotion, but the cells can glide forward or backward parallel to the long axis of the filaments. There is no morphologically distinct nucleus, and reproduction is most commonly by simple asexual cell division. These traits suggest a design similar to true bacteria. Some Cyanobacteria have internal membranes, or lamellae, along the periphery of the cells, and apparently are extensions of the external cell membrane. These apparently have a function connected with photosynthesis. The cell walls of the blue-green algae are similar to those of bacteria.
Cyanobacteria are recognized by their blue-green color, hence the name, but among the approximately 2,000 species now known, there is a wide range of colors. Most species have three pigments: chlorophyll (green), phycocyanin (blue), and phycoerythrin (red). The pigment particles are dispersed throughout the cytoplasm, especially along the peripheral lamellae, and in the sheath as well. The majority of these organisms are autotrophic and aerobic. They require no pre-formed organic matter for growth, but only oxygen, inorganic substances, and light, and photosynthesis utilizes the energy of light to build their protoplasm from the water and carbon dioxide in the air.
Cyanobacteria are agriculturally important. Activities of the blue-green algae are economically significant on several counts. First is the contribution of some species to soil fertility through the fixation of atmospheric nitrogen. For example, in the waterlogged soils of the Far East, where rice has been cultivated for centuries, an abundant bloom of blue-green algae develops, contributing to the nitrogen content of the soil and permitting good crops every year even without addition of manure or chemical fertilizers. Also, the oxygen liberated by the algae in photosynthesis may be beneficial. Artificial inoculation of soil with bluegreen algae has been successful in improving certain crops.
Blue-green algae are usually the first organisms to colonize barren rocks and arid soils. This surface growth may result in an accumulation of organic matter of such thickness that bacteria and fungi, and eventually complex plant forms, may live there. In the sea, algae are important food sources for fish and other marine life. Overgrowths of algae often develop around the intake pipes of water-supply systems and in reservoirs or cisterns of fresh water, imparting disagreeable odors. Algal blooms are likely to occur around collections of sewage-polluted water. The water consequently becomes depleted of oxygen, and fish cannot survive in it. Most of these difficulties in sanitation are controlled, however, by treatment of affected areas with copper sulfate or similar algaecides.
Summary and Conclusions
In summary, the Eubacteria, Cyanobacteria, and Archaea are tiny but very successful, judging from their ability to live in virtually every habitat on earth. Bacteria have adapted to more different living conditions than any other group of organisms. They inhabit the air, soil, and water, and exist in enormous numbers on the surfaces of virtually all plants and animals. Their diverse nutritional processes and complex genetics and cellular structure are not as different from so-called higher organisms as once believed. The Creator must have deemed bacteria important because of their ubiquity in the world, contributing to nutrient cycles and energy flow. Bacteria are also important in the body’s normal flora, and many, like E. coli, produce vitamins for the body. Bacteria are clearly made of designed structures and perform purposeful functions.
Of course, some bacteria are harmful. Certain species multiply within the human body, digesting tissues (e.g., tuberculosis) or producing disease-causing toxins (e.g., anthrax). Other bacteria infect plant crops and animal herds. Disease-causing bacteria are a global threat to all life forms in their natural state, and in the 21st century they have already been used as agents of bioterror. Bacteria have threatened human existence over the millennia with bubonic plague, typhus, tuberculosis, pneumonia, streptococcal infections, diphtheria, E. coli, and anthrax. These diseases are likely to remain until the Lord comes back and establishes a new heaven and earth.
The Genesis of Germs

Professor Alan Gillen shows that constantly mutating diseases are proof for devolution rather than evolution.
Read Online
Master Books and Dr. Alan Gillen of Liberty University have graciously granted us permission to publish selected chapters of this book online. To purchase a copy, please use the link above.
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis