

The Origin of Fleas and the Genesis of Plague
Abstract
Human history has been riddled by diseases spread by flea vectors including the bubonic plague. Recently, Madagascar has documented more than 100 cases. Fleas were part of God’s “very good” (Genesis 1:31) creation that transformed to an ectoparasitic condition after the Curse. Creation biologists continue to synthesize a working model of this conversion from environmental recyclers to parasites as a secondary state. Morphological design of the flea combined with its unique feeding ability has resulted in it being an exceptional vector for Yersinia pestis, the causative bacterium of the plague.
Introduction
Many are familiar with fleas (fig. 1) on their dogs and cats. In ages past, fleas caused plague and typhus, killing millions. Bubonic plague has started to spread again in the world, starting in Ankasina, Madagascar’s capital. As of December 2014 there have been more than 126 cases and 47 deaths. Most of the people from Madagascar, the large island off Africa, died from the bubonic form of plague. They contracted it after being bitten by a rat flea that had contracted the bacteria from the rodents.
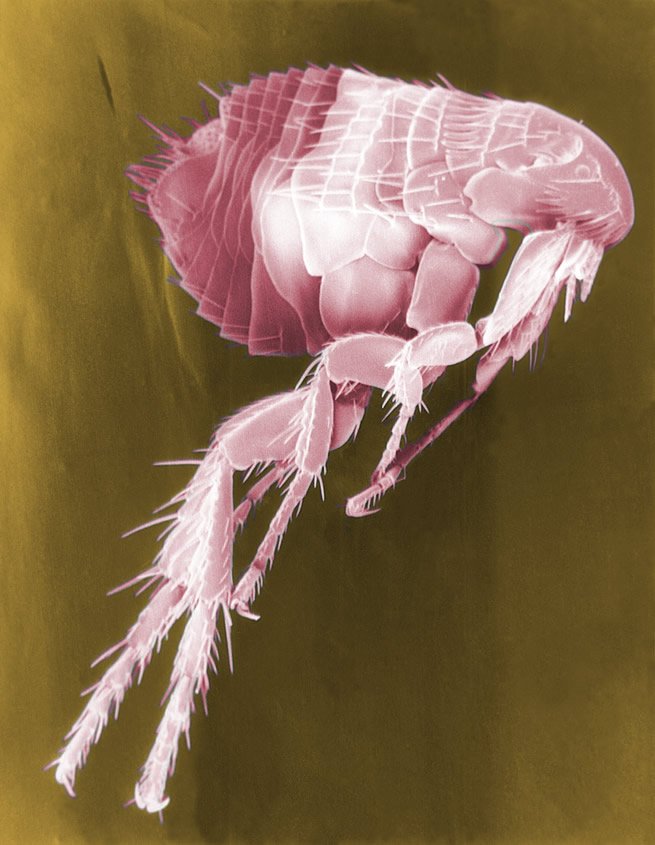
Figure 1. Scanning Electron Micrograph (False Color) of a Flea. Image reproduced from Wikimedia Commons.
Each year, the World Health Organization (WHO) receives reports of 1,000 to 2,000 cases. Between 1900 and 2010, there were 999 “confirmed or probable” cases in the US. Not many have heard of it since Ebola has captured the attention of the world. But sylvatic (occurring in wildlife) plague occurs in small numbers every year.
The Origin and “Discovery” of the Flea
Among the “creeping things” God created in Genesis 1:24–25 are countless forms of insects, including fleas.
We tend to think of them as repulsive pests, but they were created with good purposes and reveal creative diversity, design, wisdom, and the glory of God just as clearly as the fearfully and wonderfully designed human body. The ability to leap or jump is an awesome testimony of elegantly engineered, multifaceted design. They were most likely created as plant eaters to help with the recycling of leafy organic matter and detritus as flea larvae do today.
The wondrous design of the flea was first described by Robert Hooke, a Christian and creationist, in Micrographia (1665). In fact, it was his drawings of fleas using one of the earliest compound microscopes that would influence Leeuwenhoek to turn his attention to making microscopes and studying microorganisms and parasites. Hooke described and illustrated (fig. 2) a creature normally known only as a speck that bites. He listed detailed flea anatomy and illustrated how complex this little body was: “The strength and beauty of this small creature, had it no other relation at all to man, would deserve a description.” He notes the flea’s powerful jumping mechanism: “These six leggs he clitches up altogether, and when he leaps, springs them all out, and thereby exerts his whole strength at once.” And “as for the beauty of it, the Microscope manifests it to be all over adorn’d with a curiously polish’d suit of sable Armour, neatly jointed, and beset with multitudes of sharp pinns, shap’d almost like Porcupine’s Quills, or bright conical Steel-bodkins; the head is on either side beautify’d with a quick and round black eye, behind each of which also appears a small cavity, in which he seems to move to and fro a certain thin film beset with many small transparent hairs, which probably may be his ears; in the forepart of his head, between the two fore-leggs, he has two small long jointed feelers, or rather smellers . . . .”
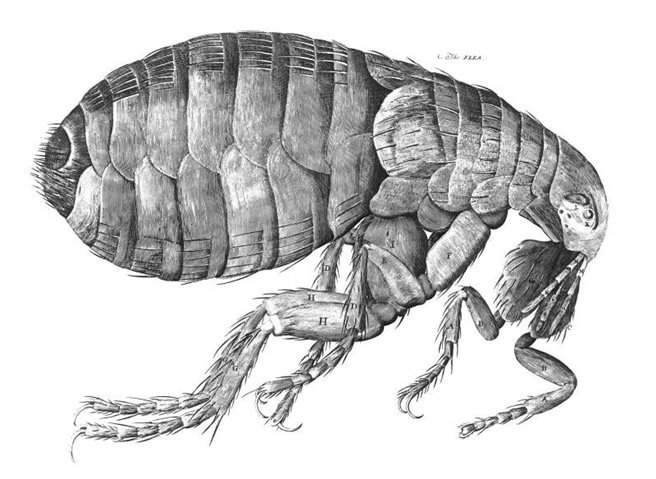
Figure 2. The Flea by Robert Hooke in Micrographia. Image reproduced from Wikimedia Commons.
This wondrous creature, its body armed with plates and spikes as described by Hooke’s words and pictures, incites appreciation, not horror. Perhaps he saw intricate planning by an ingenious Bioengineer of the flea body, especially the extraordinary hind legs. The flea is built on the basic bilateral animal model, with head, body, and limbs. It is not a collage, or a fiction, composed of random bits. Hooke’s illustration (fig. 2) stresses how its separate parts beautifully integrate into a complete, fully fashioned organism designed by God.
The Flea Kind and Unique Anatomy
There are 1,830 different kinds of fleas known throughout the world. They commonly appear on cats, dogs, and other pets. Today, we know them as ectoparasites and vectors of disease, plague, and other pestilence.
Fleas are small (1.5 to 3.3 mm long), laterally flattened wingless insects forming the order Siphonaptera (figs. 3 and 4). Fleas have well-designed hind jumping legs and mouthparts with a prominent proboscis adapted for piercing tissues (probably plants in pre-Fall era) such as the skin and sucking blood (probably plant juices in pre-Fall era). They eat plants, detritus, and organic matter as larvae and stages prior to becoming adults. The adults primarily need blood for producing eggs.
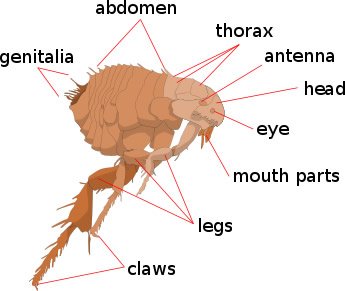
Figure 3. Basic Flea Anatomy. Image reproduced from Wikimedia Commons.
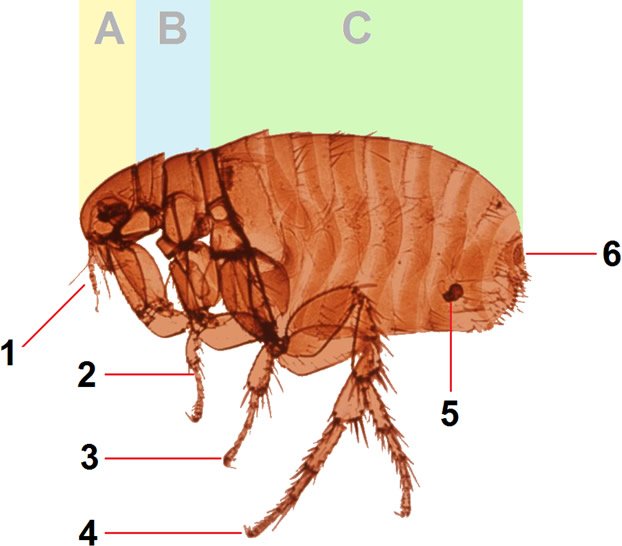
Figure 4. Advanced Flea Anatomy. Image reproduced from Wikimedia Commons.
Fleas have a unique organ called the pygidium (plural pygidia), the terminal segment of the flea designed to detect air currents. Pygidia may help in “hitchhiking” on a host (Roberts, Janovy, and Nadler, 2013). Yet, as one looks at a flea under the microscope, the most impressive feature is its exceptionally long hind legs. Most fleas are capable of vertical jumping about a foot (some fleas as high as 34 inches!), or about 150 times its own length. This is certainly worth an Olympic medal, and would be the equivalent of a human leaping up over 1,000 feet. It can jump horizontally about 13 inches, or 100 times its body length (Marquardt, Demaree, and Grieve, 2000). Lyon (2007) reports some can “long jump” horizontally up to 200 times their body length. The hind legs of fleas are linked to a zone where kinetic energy for jumping is released. The “spring” of the flea comes from an amazing “rubber-like” cross-linked protein called resilin. This substance gives fleas the capacity to easily leap upon and hitchhike with a mammal or bird host.
Most evolutionary biologists believe that fleas once had wings that were lost through time while larger hind legs evolved and that they are descendants of Mecoptera (scorpion flies). But we believe they were specially created as a distinct “kind”—designed fully formed to jump (not fly) and travel via an animal host. Fossil fleas consist mainly of modern-looking species that supposedly go back 65 million years. A possible large, non-jumping variant has been found as low as the Jurassic.1 Fossils document that fleas have always been fleas (Rothschild et al., 1973).
Phoresis
Phoresis is a symbiont carried by a host. In a sense, it is a “traveling together” of two unrelated organisms. The easiest examples to understand are pollen on a bee leg (fig. 5) or barnacles on a whale (fig. 6). Phoresis is one of the Creator’s ways to help transport many arthropods in nature to specific habitats. After the Fall and subsequent Curse, not only did arthropods become parasitic on their transport hosts (micro-predators) by eating their blood, but even communalistic bacteria within them have become pathogenic by degeneration (Gillen, 2014).

Figure 5. Phoresis. Bees Collecting Pollen. Image reproduced from Wikimedia Commons.

Figure 6. Barnacles and Whales. Image reproduced from Wikimedia Commons.
Creation biologists suggest fleas have had a phoretic relationship with other animals, having no detrimental effects on the animal host. Like the mosquito previously discussed in “The Genesis of Malaria” (Gillen and Sherwin, 2013), fleas may have utilized a protein similar to leghemoglobin (a red oxygen-binding plant hemoprotein). In today’s world, mosquitoes cannot reach the leghemoglobin found in the roots of soy plants. But was this always the case? Perhaps a similar protein once existed in an extinct nitrogen-synthesizing plant, or perhaps mosquitoes once had a way of accessing this protein found in soybeans.
Hitchin’ a Ride—Designed to Move and Disperse and Fill the Earth
Fleas probably started out as hitchhikers on small mammals to achieve dispersal. Those on dogs and cats have special enlarged combs on their back called ctenidia to attach to hairs. Ctenocephalides felis and C. canis are the most common hitchhikers, named for these unique ctenidia (fig. 7). Perhaps most fleas started this way. The most common fleas that spread plague, however, do not have prominent ctenidia: Xenopsylla cheopis (rat flea) and Pulex irritans (human flea).

Figure 7. Cat flea. Image courtesy of Alan L. Gillen.
Jumping and attaching are clearly an aid to phoresis. The compressed body of the flea enables it to move through hairs or feathers of the host, while the backwardly projecting bristles (spines) or combs serve to anchor themselves to the host’s skin like Velcro (fig. 7). Their mouthparts also aid in securely anchoring to their host. The combined quick, extraordinary jumping ability and holdfasts allow fleas to “hitch a ride” easily. This may be one method by which the Creator designed dispersal of the creeping things—to fill the earth.
Parasitism as a Secondary State
Parasitism is a type of symbiosis where the parasites exhibit a high degree of specialization and reproduce quickly and in vast numbers at the host’s expense. Classic examples of human parasitism include interactions between man and arthropods (ectoparasites). Ectoparasites are those that live on or in the skin, or attached to hairs. They include fleas, ticks, mites, and lice.
Creation biologists would suggest that parasitism is a secondary state in nature. After Creation, God cursed the earth because of man’s sin, resulting in some of the original animals and other life forms degenerating into parasites. Some have considered this relationship as “broken fellowship.” We postulate that fleas, ticks, mites, and lice may have started to be “hitchhikers” as phoretic associations.
The Origin of Bubonic Plague—From Genesis of Germs
Fleas are vectors of the bacterium Yersinia pestis, causative organism of the plague (figs. 8 and 9). Fleas bite infected mammals having Yersinia in their bloodstream. The bacteria multiply in the flea’s gut, and, when the insect in turn bites humans, the plague is transmitted. In a plague-infected flea, the bacteria forms a biofilm that eventually blocks the proventricular teeth of the digestive tract. Additional blood meals cannot pass this barrier; it is contaminated by Yersinia, and the infected blood is regurgitated into the bite.
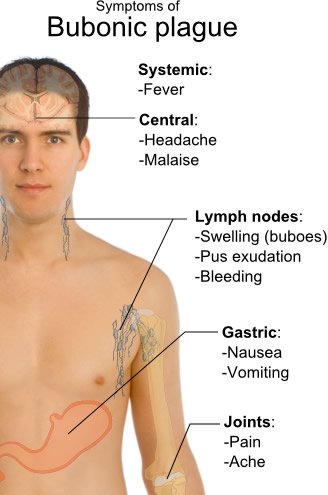
Figure 8. Symptoms of Bubonic Plague. Image reproduced from Wikimedia Commons.
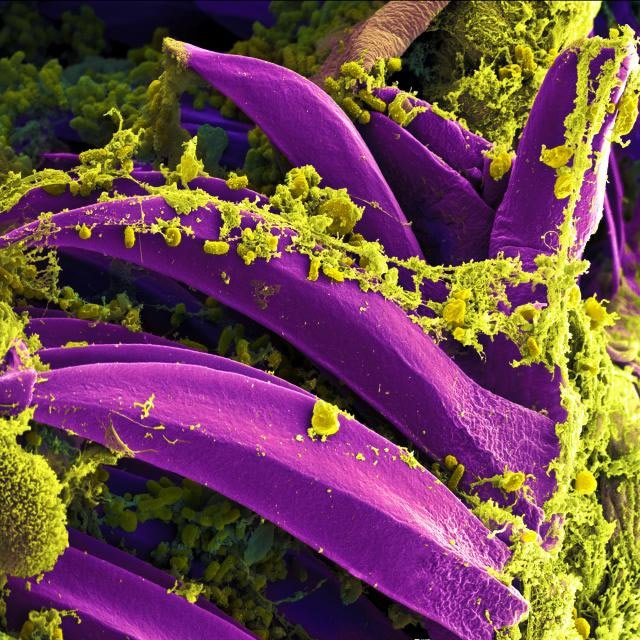
Figure 9. Scanning electron micrograph of Yersinia pestis: The Bacteria of Plague. Image reproduced from Wikimedia Commons.
A possible “good” pre-Fall function of Yersinia was to aid in digestion, not inhibit it. We postulate this because Yersinia is in the same bacteria family as E. coli (Enterobacteriaceae). E. coli and other enteric bacteria typically aid with digestion and provide vitamins for their host.
Plague occurs in more than 20 countries worldwide and is endemic in the American Southwest. Fleas on cats, squirrels, prairie dogs, and rats carry Yersinia naturally, causing sylvatic plague. Fleas that ride rodents and pass to humans to feed on our blood may carry the plague classified as bubonic, pneumonic, or septicemic.
During the 14th century, 25,000,000 people died from the plague in Europe—over one-third of the entire population. From 1347 to 1351, over 75 million people died around the world. Today, the plague commonly kills about 1 in 10 people who are infected. After the bite of an infected flea, the saliva containing Yersinia enters the bloodstream, resulting in a high fever and swelling of lymph nodes (a bubo) within a few days. In untreated cases, the bacteria infect the lungs and nervous system. Additional symptoms include headache and extreme pain (fig. 8). In some cases, the patient’s skin turns black, blue-black, or dark purple, with death from septicemia usually following. After diagnosis, patients are given antibiotics. Chances of recovery are better if diagnosed early, although some of the strains found in the 2014 Madagascar epidemic are resistant to antibiotics.
Although some forms of Yersinia (fig. 9) are harmless, other forms have devastated human populations, likely such as the plague described in Psalm 91:3–10 and in 2 Samuel 24:14–25. Perhaps the clearest evidence of the plague is recorded in 1 Samuel 6:4–19, where there is a specific reference to the tumors on people (buboes = the tumors of lymph glands) and to rats (the animal vector that carried the plague bacterium, Yersinia pestis). The biblical time frame for the plagues described in 1 Samuel was about 3,000 years ago. Interestingly, experts on plague “evolution” estimate the emergence of Y. pestis about 1,500–20,000 years ago.
Plague’s Origin Is Multifaceted
Creation biologists recognize that infectious diseases can ultimately be traced back to the decay and corruption of the original created design of microorganisms as a result of the Fall. Corruption literally means to destroy (from the Latin, corruptus). In biological terms, corruption is a loss of genetic information or a deleterious change to genetic material; whether an insertion or a deletion, the result is damage to the original code. The origin of the pathogenicity (disease-causing capability) of bacteria such as in Y. pestis is both complex and multifaceted, and it may be explained by a combination of genes that were lost, added, and moved. The story of Yersinia’s degeneration into the plague pathogen may serve as a model of “fast” genomic decay and corruption (Gillen and Sherwin, 2006).
It appears the beginning of pathogenicity in the genus Yersina started with a net loss of chromosomal DNA from its original “kind.” Later, there were minor additions of plasmid DNA (extrachromosomal DNA) as well as viral and other bacterial DNA. A few plasmid genes for toxins have been acquired from other existing species, but many chromosomal genes were likely lost. It takes only a few such gene changes to produce a new and extremely infectious variant, so it may have taken only centuries or a few thousand years to produce the current bubonic plague strain that has existed for about the last 500 years.
Researchers hypothesize that key chromosomal genes (i.e., involved in metabolic pathways) were lost in the change from a soil-inhabiting Yersinia to a pathogenic Yersinia species. Pathogenic Yersiniae have seemingly lost the genetic expression of numerous genes (about 149). Of the genes lost, 58 are the result of frameshift mutations, 32 have undergone deletions, and the rest are nonsense mutations—all of which prevented pre-existing genes from being normally expressed.
An important feature of the Y. pestis genome is the presence of pseudogenes. Biologists believe these genes reflect a loss of structural information and function. Dr. Wren (2003), the leading plague expert, suggests that the genes lost in Y. pestis affected bioenergetic functions. This reduction of metabolic pathways could have allowed the bacterium to conserve energy. The newly emerged strains (variants) are streamlined and could have contributed to the development of pathogenicity (i.e., plague traits) due to the genes they lack. The absence of important bioenergetic genes is believed by geneticists to be a hallmark of genomic decay.
Genes Added and Moved
The corruption by three genes in particular of a relatively benign ancestor of Y. pestis may have played a key role in the emergence of bubonic plague. A report by Hinnebusch and colleagues (2002), a team of plague experts at the NIH, suggests that the acquisition of two plasmid genes from other bacteria in recent times changed the fairly harmless Y. pseudotuberculosis, which caused mild food poisoning, to the agent of the “Black Death.” A third gene (carried on plasmid pMT1) produces murine toxin, an enzyme required for the initial survival of Y. pestis bacilli in the flea midgut. By acquiring this gene from another organism, Y. pestis made a crucial shift in its host range. The bacterium now could survive in fleas, and it devolved to rely on its blood-feeding host for transmission. This is just another example of the flexibility of microbes and how they sometimes repackage themselves into more dangerous agents of infectious disease via the corruption of genetic information.
This last corruption was one that distinguishes Yersinia pestis from all closely related, more benign bacteria such as Y. pseudotuberculosis and other Yersinia species. As Y. pestis adapted to rely on its new blood-feeding host for transmission, our research with media containing blood has shown that the emergence of pathogenic bacterial strains would have been favored. It appears that these minor plasmid additions were the last changes made in an otherwise long series of genetic losses in Y. pseudotuberculosis’ chromosome.
One pathogenicity island (on plasmid pMT1) was probably acquired by Yersinia pestis from a different bacterium. As with the others, these genes were acquired through lateral gene transfer and inserted into the bacteria’s DNA. It produced a message corrupted from its original “very good” purpose that gave bacteria a new niche in the foregut. Y. pseudotuberculosis that lacks the chromosomal hms locus gene harmlessly inhabits the flea midgut. Plague bacilli, by contrast, have this inserted gene. Free from their original control, the bacteria can now migrate from the midgut to the foregut, forming a plug of packed plague bacilli that is passed on to the victim when the flea feeds (Gillen and Sherwin, 2006; Gillen, 2014).
Summary and Conclusions
The wondrous “very good” design of fleas may be effaced, but not entirely erased. The likely purpose of fleas is to aid in the decomposition of decaying matter and to enrich the soil. In their early life stages, they feed on detritus, dead insects, and decaying plant matter. Their extraordinary jumping ability probably allowed them to hitchhike on animals and disperse throughout the world. But rather than always being a pest that evolved from a different insect, fleas are a wonder of God’s creation that have undergone a conversion from the Curse. This is seen in a type of symbiosis called external parasitism, an interaction between man and arthropod. It is suggested that fleas, ticks, mites, and lice all started as these “hitchhikers” in a phoretic association, transporting these arthropods to specific habitats. Although fleas may have started as mere phoretic travelers, at some point in time they became ectoparasites, then vectors. And even evolutionary biologists see their change to vectors of diseases like bubonic plague happening only several thousand years ago.
Fast Facts
- Communalistic bacteria associated with arthropods such as fleas became pathogens by degeneration.
- Yersinia pestis is the causative bacterium of the plague that occurs in more than 20 countries worldwide.
- The sylvatic plague occurs in wildlife while fleas that ride rodents pass bubonic, pneumonic, or septicemic plague to humans when they feed on our blood.
- When patients are diagnosed early, they have better chances of recovering.
- Some of the strains of the plague currently found on Madagascar are resistant to antibiotics.
- Bubonic plague (the “Black Death”) killed one-fourth of Europe’s population in the 1300s.
- A biblical record of the plague is recorded in 1 Samuel 6:4–19, where there is a reference to the tumors on people and to rats. Secular researchers estimate the emergence of Y. pestis about 1,500–20,000 years ago—within the biblical timeline.
- Creation scientists maintain a number of infectious diseases can be explained by the corruption of the original created design of microorganisms.
- Corruption is a loss of genetic information or an addition of genetic material that damages the original purpose.
The origin of Y. pestis is complex, and it may be explained by a combination of genes that were lost, added and moved, serving as a model of “fast” genomic decay and corruption. This is merely one way an organism can adapt to live in this fallen world, not an example of a pathway for molecules-to-man evolution. Indeed, researchers suggest key chromosomal genes were lost in the change from a soil inhabiting Yersinia type to a pathogenic Yersinia species. Research continues as creation biologists investigate the origin of parasitism and the genesis of disease, while giving honor to the original designs and still-sustaining omnipotence of the Creator (Psalm 139:17; Colossians 1:16–17; Acts 17:25).
References
Gillen, A. L. 2014. The Genesis of Germs: Disease and the Coming Plagues in a Fallen World. Green Forest, Arkansas: Master Books.
Gillen, A. and Sherwin, F. 2006. “The Origin of Bubonic Plague.” Journal of Creation 20 (1): 7–8.
Gillen, A. L. and Sherwin, F. 2013. “The Genesis of Malaria: The Origin of Mosquitoes and their Protistan Cargo, Plasmodium falciparum.” Answers in Genesis. https://answersingenesis.org/biology/disease/the-genesis-of-malaria/.
Hinnebusch, B.J., Rudolph, A.E., Cherepanov, P., Dixon, J.E., Schwan, T.G., and Forsberg, A., 2002. “Role of Yersinia Murine Toxin in Survival of Yersinia pestis in the Midgut of the Flea Vector.” Science 296: 733–735.
Hooke, R. 1665. Micrographia or Some Physiological Descriptions of Minute Bodies. New York: Cosimo Classics.
Lyon, W. F. 2007. “Fleas.” Ohio State University Extension Fact Sheet, Entomology page. http://ohioline.osu.edu/hyg-fact/2000/2081.html (accessed December 30, 2008).
Marquardt, W. C., R. S. Demaree, and R. B. Grieve. 2000. Parasitology and Vector Biology, 2nd edition. San Diego, California: Harcourt Academic Press.
Roberts, L. S., J. Janovy, Jr., and S. Nadler. 2013. Schmidt and Roberts’ Foundations of Parasitology, 9th edition. Boston, Massachusetts: WCB McGraw-Hill.
Rothschild, M. Y. Schlein, K. Parker, C. Neville and S. Sternberg. 1973. “The Flying Leap of the Flea.” Scientific American 229 (5): 92–100.
Tortora, G. J., B. R. Funke, and C. L. Case. 2013. Microbiology, An Introduction,11th edition. San Francisco, California: Pearson Benjamin/Cummings Pub. Co.
Wren, B.W., 2003. “The Yersiniae—A Model Genus to Study the Rapid Evolution of Bacterial Pathogens.” Nature Reviews Microbiology 1: 55–64.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_Genitalia_-_Clusters_of_Barnacles_surround_the_Genitalia_-_3_July_2014.jpg){kind=link}
{kind=link}
{kind=link}
About the author: Dr. Alan L. Gillen is Professor of Biology at the Department of Biology and Chemistry, Liberty University, 1971 University Boulevard, Lynchburg, VA 24502
Footnotes
- Elizabeth Mitchell, “Paleo-Pest Said to Demonstrate How a Flea Evolved into a Flea,” Answers in Genesis, July 15, 2013, https://answersingenesis.org/natural-selection/adaptation/paleo-pest-said-to-demonstrate-how-a-flea-evolved-into-a-flea/.
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis