

Walking Whales on Noah’s Ark?
Could Whale Evolution Fit Within Biblical Creation?
For most animals that needed to be taken on Noah’s ark, the criteria seem rather straightforward. They needed to be terrestrial (or what is termed at the Ark Encounter, land-dependent), air-breathing, and either flying creatures or crawling/hopping/walking/slithering creatures (as recounted by the creatures which perished outside the ark—per Genesis 7:21–23). That sums up the human passengers as well as the vast majority of reptiles, amphibians, and mammals which were on the ark. If there is any “fuzzy” area here, it is definitely the term terrestrial (or land-dependent). What about semiaquatic animals? Could most of them survive outside the ark, or did they need to be on the ark? And what of animals which are now completely freshwater- or sea-dwelling but may have speciated out from more terrestrial members of their kind?
Semiaquatic Animals Included on or Excluded from the Ark
Otters are listed as being onboard Noah’s ark in our “Mammalian Ark Kinds” article. Yet marine otters are mostly aquatic so may have had no need to be on the ark; however, it was determined that they are of the Mustelidae (weasel/otter/badger) kind, which are mostly terrestrial, so marine otters likely have developed their aquatic lifestyle post-flood. Many birds are also semiaquatic (or even mostly aquatic), yet all the bird kinds are listed as ark passengers. The same can be said for some turtle genera and most crocodilians, which are mostly aquatic creatures, yet they were also included as being on the ark. The same holds true for sea kraits (a type of sea snake) because some species within that genus (Laticauda) are more terrestrial.
Many other semiaquatic animals, however, are not listed as being ark occupants, as they either only come onto land to lay eggs (sea turtles) or give birth, mate, and occasionally bask (seals, sea lions, and walruses). And then there are fish that spend considerable amounts of time on land, like the Pacific leaping blenny, lungfish, and mudskippers—none of which are listed as ark passengers even though the lungfish definitely is “air breathing” and frequently “lives” on land.
Semiaquatic Whales?
If whales existed as obligate marine creatures before the flood, why are there no fossils of them in flood strata?
So how does this discussion of semiaquatic animals apply to whales, which are fully aquatic? Well for starters, they are air-breathing mammals. And there are some young-earth creationists (YECs) who believe that, like the marine otter, whales come from a biblical kind that was terrestrial or semiaquatic at the time of the flood. If the marine otter can adapt to an aquatic lifestyle after the flood (which we would agree with), why couldn’t whales? Another aspect that drives this “walking whale” concept is that the fossils of all modern cetaceans (both baleen and toothed whales) are found in Upper Eocene (if late archaeocetes like Basilosaurus are included) to Oligocene (modern toothed and baleen whales) and higher strata, which are viewed by many YECs as post-flood. If whales existed as obligate marine creatures before the flood, why are there no fossils of them in flood strata?
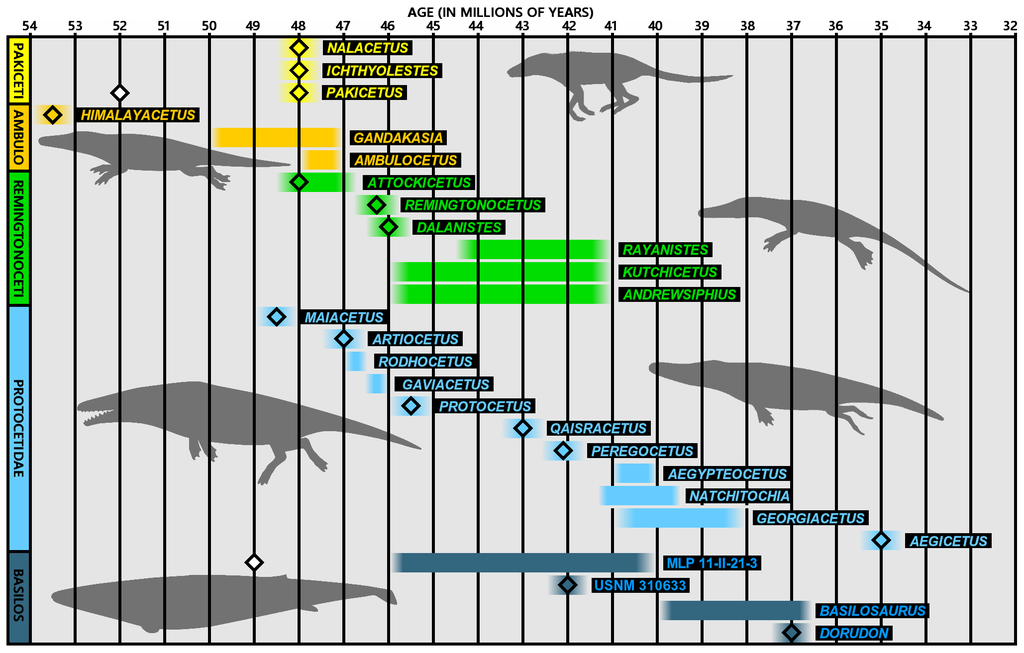
Fig.1 Sneaking stoat 2, CC BY-SA 4.0, via Wikimedia Commons
This has caused some researchers to conclude that perhaps some of the creatures proposed by evolutionary biologists to be evolutionary ancestors of whales were in fact semiaquatic whales, which diversified into modern whales post-flood. Geologist Kurt Wise has summed up this thought in a few articles and book chapters (direct quotes below).
Finally, some of the animals which are aquatic or marine today may not have been aquatic at the time of the Flood. The marine and sea otters, for example, are members of the mustelid (weasel) family and their aquatic character is likely to have been revealed after the Flood. The whales might turn out to be another example. Only when including the legged archaeocetes (and/or possibly the terrestrial order Acreodi) do the whales have a fossil record continuous with the Flood. Vestigial legs and hips in modern whales confirm legged ancestors of the whales existed only a short time ago. It is possible that the purely marine cetaceans of the present were derived from semi-aquatic or even terrestrial ancestors on the ark.1
Some of the changes that have occurred among organisms seem to be evidenced in vestigial structures (feathers that had a strong function in the past but now seem to have reduced function or no function at all) and genetic throwbacks (past structures that appear spontaneously in a small percentage of offspring in the present). Hip and leg bones that appear in some foetal sperm whales, for example, are vestigial structures. They suggest that modern whales might be descendants of whales in the past that had hind limbs.2
At this point in time, the largest challenge from the stratomorphic intermediate record appears to this author to come from the fossil record of the whales. There is a strong stratigraphic series of archaeocete genera claimed by Gingerich (Ambulocetus, Rhodocetus, and Prozeuglodon [or the similar-aged Basilosaurus] followed on the one hand by modern mysticetes, and on the other hand by the family Squalodontidae and then modern odontocetes. That same series is also a morphological series: Ambulocetus with the largest hind legs; Rhodocetus with hindlegs one third smaller; Prozeuglodon with 6 inch hindlegs; and the remaining whales with virtually no to no hind legs: toothed mysticetes before non-toothed baleen whales; the squalodontid odontocetes with telescoped skull but triangular teeth; and the modern odontocetes with telescoped skulls and conical teeth. This series of fossils is thus a very powerful stratomorphic series. Because the land mammal-to-whale transition (theorized by macroevolutionary theory and evidenced by the fossil record) is a land-to-sea transition, the relative order of land mammals, archaeocetes, and modern whales is not explainable in the conventional Flood geology method (transgressing Flood waters). Furthermore, whale fossils are only known in Cenozoic (and thus post-Flood) sediments. This seems to run counter to the intuitive expectation that the whales should have been found in or even throughout Flood sediments. At present creation theory has no good explanation for the fossil record of whales.3
During the development of some sperm whales the fetus develops portions of hind limb and pelvic bones and subsequently resorbs them. Other modern cetaceans have other bones in embryology and sometimes adulthood which suggest hind limb vestiges. This suggests that modern cetaceans have latent genetic information for the development of small hind limbs. Could this mean that Basilosaurus, for example, (which has hind limbs and pelvis) is actually an early post-Flood representative of some cetacean baramin and that modern cetaceans of that baramin are actually descendant from it — or something similar to it?4
Examining the Wise (and Otherwise) Arguments
Vestigial Hind Limbs
Let’s begin by examining the “vestigial limbs” found on whales and dolphins. It has certainly been documented in scientific literature, as well as anecdotal accounts from fishermen, that on rare occasions small rear flippers have been found on dolphins and small bones have been found on the backs of some whales. Counting both the scientific and anecdotal accounts, starting with Roy Chapman Andrews’ 1921 report of a 1919 humpback whale find off Vancouver island all the way to the 2006 find of a dolphin with two rear fins, we have about a century of observed cases. Yet for all this time, we have approximately 10 cases. One case involved a herd of 450 blue-white (or striped) dolphins, caught by Japanese fisherman off the eastern coast of Izu Peninsula. Out of the 450 dolphins, only one had what was described as “rudimentary hind limbs.” Yet no mention of bones was in the account, so these were likely a pair of rear flippers. All other accounts of vestigial hind limbs are similar singletons, but some have been documented and photographed showing bones in right whales, sperm whales, and humpback whales. Most of these bones are ridiculously small, often just several centimeters, and bear no resemblance to femurs and tibia (although evolutionary biologists frequently make that claim).5
Rather than proof of an evolutionary lineage, they are proof that we live in a cursed world.
Using the example of the 450 dolphins above, why was only one found with “vestigial” hind limbs? And out of the many whales killed for food, accidentally killed, or beached over the past 100 years, why have so few “vestigial” limbs been reported? Paleontologist Roy Chapman Andrews stated this in 1921 after examining the “hind leg bones” of a female humpback whale: “After studying the material and discussing it with various scientists, I have come to the conclusion that the protrusions actually do represent vestigial hind limbs and show a remarkable reversion to the primitive quadrupedal condition.”6 If there exists a vestigial organ or set of bones on an animal, shouldn’t it be much more pervasive in specimens?7 The scarceness of these bones (or rear flippers) indicates they are much more likely the result of mutation than a genetic throwback. We occasionally see five-legged pigs8 and cows,9 yet these are never (nor should they be) claimed as vestigial limbs of an alleged six-legged porcine or bovine ancestor. Rather than proof of an evolutionary lineage, they are proof that we live in a cursed world.
The Fossil Record of True Whales
Dr. Wise does bring up a cogent point in that there are no true whale fossils found before the late Eocene, which is viewed by many creation geologists as post-flood. The argument goes that since we have good fossil representation of marine reptiles like plesiosaurs and mosasaurs from the flood, why not whales if they existed as obligate marine mammals and presumably inhabited the same ecological zones (deep-water environments)? One explanation is fairly straightforward: whales have blubber, whereas most marine reptiles do not, which means whales and dolphins would have been more likely to float upon dying and not be buried, thus making it more likely they were scavenged and not fossilized. Second, the flood would have created highly turbulent and toxic conditions in the oceans due to underwater volcanos, turbulent and mud-laden waters, etc. All sea life with gills (fish, including large sharks) would have been much more likely to succumb to water toxicity or have their gills covered in sediment and asphyxiate. Whales could always surface and breathe in oxygen, making them much more likely to survive the flood. Marine reptiles (which also breathe air and must surface) likely could not stay submerged very long. The average time between breaths for sea snakes, for example is 20–30 minutes.10 Whales on the other hand can routinely go 90 minutes, and one dive was recorded at 222 minutes.11 Third, do we really know that whales and marine reptiles shared the same ecological zones in the preflood world? In reality, most creation models postulate shallow oceans, which means the more likely “kill zones” for marine life during the flood were near shore vs. offshore—which would lead, again, to less representation of large sea mammals in the fossil record.
Also, while Dr. Wise accepts the K-Pg boundary as the post-flood boundary, his argument for whales would likewise need to be stretched to include pinnipeds (seals, sea lions, and walruses), whose fossil record only goes back to the late Oligocene and early Miocene (Enaliarctos and Puijila).12 And yes, Wise does believe that most semiaquatic mammals might have been on the ark or might have been represented by fully terrestrial ancestors as well (in contrast to the Ark Encounter’s stance).
Mammals such as otters, beavers, muskrats and even seals may have may have been able to survive on the ark without having to live in the water. Finally, some of the animals which are aquatic or marine today may not have been aquatic at the time of the Flood. The marine and sea otters, for example are members of the mustelid (weasel) family and their aquatic character is likely to have been revealed after the Flood. The whales might turn out to be another example. Only when including the legged archeocetes (and/or possibly the terrestrial suborder Acreodi) do the whales have a fossil record continuous with the Flood.13
But let’s consider the coelacanth, which was supposedly extinct since the end of the Cretaceous (usually the latest fossils are dated to about 80–70 mya), and yet they still exist today. Likewise, Lavesia, a genus of atelestid fly originally discovered in 1999 as a fossil in amber and believed to have gone extinct over 100 million years ago, was found to be a living species found in Namibia in 2010. And Monoplacophora, a class of mollusks, was believed to have gone extinct in the middle Devonian Period until living members were discovered in deep water off Costa Rica in 1952. All of these would be considered non-ark occupants which clearly survived the flood yet left no fossil record in late-flood (Cretaceous) or post-flood rocks.
Biblical Arguments Against the Land-to-Sea Whale Fossil Series
The term used for “created kind” in creation biology is baramin, which was derived by creation scientists from two Hebrew words: bara (translated as “create” in Genesis 1:1, 21, and 27) and min (translated as “kind” in Genesis 1:21 and 24–25, when God created the sea, winged, and land creatures on days five and six and in Genesis 6:20 and 7:14 to refer to land animals and flying creatures that Noah brought on board the ark). Of particular interest to this discussion is Genesis 1:21’s account of day five: “So God created the great sea creatures and every living creature that moves, with which the waters swarm, according to their kinds, and every winged bird according to its kind. And God saw that it was good.” Contrast this with the day-six creatures God created: “And God said, ‘Let the earth bring forth living creatures according to their kinds—livestock and creeping things and beasts of the earth according to their kinds.’ And it was so. And God made the beasts of the earth according to their kinds and the livestock according to their kinds, and everything that creeps on the ground according to its kind. And God saw that it was good” (Genesis 1:24–25).
It’s doubtful that there are many (if any) creationists out there who would argue that lungfish were created on day six as a “creeping kind.” However, why not argue that they were created on day five as sea creatures, yet by the time of the flood had become a terrestrial kind and had to be on Noah’s ark since they breathe through their lungs and can drown?14 They have small limbs (which makes them semiaquatic), and “the larvae have external gills that are reabsorbed during their metamorphosis into fully developed lungfish.”15 (Any of this sound familiar?) So why not lungfish as ark occupants? I think that we can see that they are still “fish” despite their adaptability to semiaquatic or drought conditions.
But more importantly, we also see that the aquatic animal kinds were created on day five for a reason: to “be fruitful and multiply and fill the waters in the seas” (Genesis 1:22). Notice also that after creating the land creatures (and before he created man), God saw that it was good (Genesis 1:25). If they were created as terrestrial or land-dependent semiaquatic creatures and were declared “good” in their design, why and how would they move to the sea with their designed-for-land features? Most YECs accept that there are kind boundaries that cannot be crossed, and the day five and six creatures certainly fit into this biblical-kind exclusion. In other words, a day six created kind could not become a day five creature, or vice versa.
Developmental Arguments Against the Land-to-Sea Whale Fossil Series
As PhD marine biologist Robert Carter has said,
The evolutionary ancestors of whales could not nurse their young underwater. Their breathing and eating systems were still connected. They would have had no need for those strange mouth plates that baleen whales use to filter out krill from the water. They would not have had the ability to echolocate. There are entire suits of morphological and behavioural changes that are necessary to explain whales.16
Creation geologist John Woodmorappe discusses the tail fluke of whales and says in reference to a study on whale fluke evolution, “It is not only the limbs, but also the tail, which supposedly underwent extensive modifications in order to convert a terrestrial creature into an aquatic one. Entirely omitted in the National Geographic article is the fact that, owing partly to preservation problems, there is a lack of intermediates between tails and flukes.”17
Dr. Brian Thomas of ICR covered some additional features of rorqual whales (blue and minke whales) that show there are too many diverse and finely tuned irreducible complexities in blue and minke whales for them to have evolved all at once: “The precisely arranged body parts of rorqual whales were all required to coexist simultaneously for them to eat and survive, confounding whale evolution and confirming Genesis 1:21.”18 In another article, Thomas stated this in more detail:
A subset of baleen, rorqual whales eat through a process known as “lunge feeding” that requires a long list of fully formed features. One of those unique traits, essential for the whale’s feeding, involves the mouth and the jaw—the accordion-like skin folds on the throat greatly expand when they swallow huge mouthfuls of prey-laden water.
No combination of natural processes could have organized rorqual whale features, even considering toothed whales as possible evolutionary precursors. All the necessary traits were required for survival in the beginning, so they must all have popped into existence by a miraculous creation event.
In the journal Nature, American and Canadian scientists described a previously unknown sensory organ situated in the front and center of the whale’s lower jawbone, where the bone is split into left and right halves. The organ measures and informs the brain about the resistance force upon the whale’s gaping mouth when lunge feeding. The unique organ also detects “dynamic rotation of the jaws during mouth opening and closure,” according to the report. In other words, without this sensory organ whales would not know how much force is too much when lunging through water—they could fatally damage themselves without this key sensory and data coordination device.
And according to the study authors, baleen whales need all of the following parts linked in precise proportions in order to eat: comb-like baleen to filter out food; expandable, accordion-like “ventral groove blubber” with cartilaginous support bars; the newly discovered sensory organ; a split jaw that is loosely connected to the skull; and tactile organs, “vibrissae,” along the chin that sense prey.19
No Walking Whales
Another issue with whales “walking” off the ark is that it implies that God’s categories of land and sea creatures can be easily and readily blurred and that God’s good design of land creatures (if they indeed later became sea creatures) was not “good enough” for them to stay land creatures.
Whales were created on day five of creation week as fully aquatic creatures. They were commanded to fill the seas and did not evolve from or speciate out from terrestrial or semiaquatic animals before or after the flood. The level of change goes way beyond inter-baraminic generation and speciation. It would require a complete body-plan overhaul and several adaptations arriving at exactly the same time for the creature to survive. And “evolution” (or YEE [young-earth evolution] hyper-fast speciation) would have to do this twice because of the vast differences in baleen and toothed whales. And in evolutionary timeframes, there are only tens of millions of years to go from terrestrial to semiaquatic to obligate aquatic creatures (which by evolutionary standards is lightning fast).
But even more disturbing in this line of thought from a creation perspective is that it overemphasizes the evolutionary “whale ancestors” in the fossil record (alleged semiaquatic whales), or the lack of one for Mysticeti and Odontoceti, prior to the late Eocene. As noted above, there are plausible models for the lack of whale fossils before the late Eocene/early Oligocene.20
Another issue with whales “walking” off the ark is that it implies that God’s categories of land and sea creatures can be easily and readily blurred and that God’s good design of land creatures (if they indeed later became sea creatures) was not “good enough” for them to stay land creatures. While some may point out that the pinnipeds and sea otters are in the same category, they are not, nor would they ever be, described as obligate sea creatures. They still mate and bear live young on land (although sea otters can and do sometimes give birth in the water) and spend considerable time basking, socializing, and even fighting on land. Sea otters have also been noticed going onto land to rest, groom, or nurse.
Finally, falling back on the outdated “ontogeny recapitulates phylogeny” argument to argue that a few individual cetaceans that have “limb buds” which get reabsorbed during early embryonic stages as proof of either an evolutionary ancestor or a terrestrial or semiaquatic land creature lineage is lamentable. As Dr. Jake Hebert has said,
YEEs tend to play up supposed evidence for evolution while playing down evidence for creation. Is this because the evidence is truly equivocal and we creationists have been overstating our position? Or is it because YEEs are philosophically opposed even to the possibility that the evidence could strongly favor creation? . . .
Biblical creationists cannot afford to get this wrong. Affirming a recent creation and a global flood is meaningless if one claims that evidence for these things is ambiguous or equivocal. Biblical creation is true, and the scientific and historical evidence strongly confirms this.21



Footnotes
- K. P. Wise, “Mammal Kinds—how many were on the Ark?” in Genesis Kinds: Creationism and the Origin of Species, eds. T. C. Wood and P. A. Garner (Eugene, Oregon: Wipf & Stock, 2009), 143.
- K. P. Wise, Faith Form and Time (Nashville, Tennessee: Broadman & Holman 2002), 219.
- K. P. Wise, “Towards a Creationist Understanding of ‘Transitional Forms,’” Creation Ex Nihilo Technical Journal 9, no. 2 (1995): 216–222.
- Wise, “Towards a Creationist Understanding of ‘Transitional Forms.’”
- Ernest C. Conrad, “True Vestigial Structures in Whales and Dolphins,” Creation/Evolution Journal 3, no. 4 (Fall 1982), https://ncse.ngo/true-vestigial-structures-whales-and-dolphins.
- Conrad, “True Vestigial Structures.”
- Think of the “vestigial organ” argument for humans. What two are mentioned the most times? The appendix and the tonsils. And how many humans have these “vestiges”? Almost 100%. Why should “vestigial hind legs” in whales be so-named if less than 1% exhibit them?
- Rob Leigh, “Pictures: pig born with five legs in China,” Mirror, updated January 28, 2012, https://www.mirror.co.uk/news/weird-news/pictures-pig-born-with-five-legs-324190.
- Kendall Davis, “5-legged cow captures visitors’ attention at Danville-Pittsylvania County Fair,” WDBJ7, September 19, 2019, https://www.wdbj7.com/content/news/5-legged-cow-captures-visitors-attention-at-the-Danville-Pittsylvania-County-Fair-560810541.html.
- “Sea snakes,” Marine Education Society of Australasia, accessed March 9, 2023, http://www.mesa.edu.au/seaweek2004/discovery15.asp.
- “How long can whales hold their breath?” Quick question, Natural History Museum, accessed March 9, 2023, https://www.nhm.ac.uk/discover/quick-questions/how-long-can-whales-hold-their-breath.html#:~:text=The%20longest%20ever%20recorded%20dive,to%20the%20surface%20to%20breathe.
- Irina Koretsky, L. G. Barnes, and Sulman Rahmat, “Re-Evaluation of Morphological Characters Questions Current Views of Pinniped Origins,” Vestnik Zoologii 50, no. 4 (2016): 327–354, https://www.researchgate.net/publication/309618340_Re-Evaluation_of_Morphological_Characters_Questions_Current_Views_of_Pinniped_Origins.
- Wise, “Mammal Kinds,” 143.
- “Introduction to the Dipnoi,” UCMP Berkeley, accessed March 9, 2023, https://ucmp.berkeley.edu/vertebrates/sarco/dipnoi.html#:~:text=The%20Dipnoi%20are%20a%20group,absorbs%20oxygen%20and%20removes%20wastes.
- “African lungfish,” Oregon Zoo, accessed March 9, 2023, https://www.oregonzoo.org/animals/african-lungfish.
- Robert Carter, “Species were designed to change, part 2: Speciation and the limits of change,” Creation Ministries International, July 22, 2021, accessed March 2, 2023, https://creation.com/species-designed-to-change-part-2.
- John Woodmorappe, “Walking Wales, Nested Hierarchies, and Chimeras: Do They Exist?” Answers in Genesis, April 1, 2022, https://answersingenesis.org/missing-links/walking-whales-nested-hierarchies-and-chimeras-do-they-exist/.
- Brian Thomas, “Pastor Became Atheist. Why?” ICR, August 8, 2012, https://www.icr.org/article/6935.
- Brian Thomas, “Organ Discovery Shows Why Whales Didn't Evolve,” ICR, June 22, 2012, https://www.icr.org/article/organ-discovery-shows-why-whales-didnt.
- Some YEC organizations believe that the post-flood boundary is much higher, near the Miocene-Pliocene boundary or even higher at the Neogene-Quaternary boundary. While this would solve the “whale flood fossil problem,” it has some difficulties with mineral- and ore-weathering patterns which would take decades to form and thus were likely caused after the flood, placing the post-flood boundary much lower in the rock record toward the K-Pg boundary. As Dr. Andrew Snelling states, “The relative ages of supergene minerals thus favor a placement at the K-Pg boundary with a relative age of 66 Ma, the Flood/post-Flood boundary favored by many Flood geologists” (Andrew A. Snelling, “Locating the Flood/post-Flood Boundary Using the Relative Dating of the Weathering of Ore Deposits,” The Proceedings of the International Conference on Creationism 8, no. 46 [2018]: 553–566, https://doi.org/10.15385/jpicc.2018.8.1.40).
- Jake Hebert, “Is Creation Evidence Ambiguous?” ICR, July 31, 2020, https://www.icr.org/article/is-creation-evidence-ambiguous/.


{kind=link}
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis