

Life’s Hidden Marvel
An engineering feat that far surpasses landing a man on the moon is taking place all around us.
Developmental biology is one of the most awesome biological disciplines to display the glory of God and the immeasurable value of human life.
The sheer scale of this developmental process and its output is a witness to God’s wisdom and power.
Think about this. You might be 5'10" today, but you were once a mere 100 microns in diameter, smaller than the dot above an i. Such astronomical growth is comparable to a marble growing to the width of a football field—a 10,000-fold increase! Consider further, your body is composed of tens of trillions of cells (about 10,000 times earth’s population), all originating from that tiny cell created at conception. Your more than 200 cell types came from that cell containing all the information necessary to direct the development and organization of the tens of trillions of cells into integrated systems so you could design skyscrapers, compose symphonies, perform athletic feats of gymnastics and flying fighter jets, speak multiple languages, and do mathematics that would put man on the moon. The sheer scale of this developmental process and its output is a witness to God’s wisdom and power.
My personal relationship with Christ the Creator has been profoundly impacted through my study of developmental biology. As I consider the complex genetic regulatory networks that control the migration of cells to their correct positions and precisely timed cell maturation and specialization, and when I consider that these immature tissues carry out functions for embryonic life at each stage, it evokes worship and fills my heart with praise to God. His infinite attention to detail in making my body assures me he is superintending each detail of my daily life. What a comfort! My devotional experience from in-depth study of development can be summarized in the antiphonal structure of Psalm 139:13–16: God’s work . . . our praise . . . God’s work . . . our trust in his care.
In Scripture, the heart is a central theme: “You shall love the LORD your God with all your heart”; “unite my heart to fear Your name”; “keep your heart with all vigilance, for from it flow the springs of life”; “with the heart one believes and is justified” (Deuteronomy 6:5; Psalm 86:11; Proverbs 4:23; Romans 10:10). Given this, let us examine some wonders of the heart’s developmental biology.
The First Organ
Despite what one might think, neither the brain, the skeleton, nor skin is first to develop in the embryo. Rather, it is the heart. The very first organ our Creator caused to form in each individual is a heart. It brings immediately to my mind the Scripture about the first (most preeminent) commandment and its corollary, according to Jesus Christ, to love God supremely with all our hearts and to love our neighbor as ourselves (Mark 12:29–31).
From a functional perspective of embryonic development, it makes sense that the heart would form first. The growing number of cells must be kept alive, including those that give rise to future neurons, osteocytes, and cells of the epidermis. The heart is needed to pump life-giving blood that delivers oxygen and nutrients and removes waste products from the embryonic cells. Thus, the circulatory system—heart, blood, and blood vessels—is the first organ system.
Making an Embryonic Heart
Not only the first organ to develop, the heart also shows a unique developmental process not seen with the other major organs. In the early embryo (in humans, less than three weeks old), the heart begins to develop bilaterally, from two separate regions (called a heart field) located on either side of the embryonic midline. Simultaneously and independently, the heart field cells self-assemble into two separate tubes. These bilateral tubes must move in the correct three-dimensional direction and find the embryonic midline. The tubes have the ability to detect local migratory cues, pointing the way to the midline where they then fuse with the corresponding tube from the other side. It is now a common heart tube and eventually will become the left ventricle, the largest and “power” chamber of the heart. Uniting two tubes from separate regions of the embryo is an astounding feat of genetically regulated cell communication and migration—all to form just one of the four chambers of the heart!
Meanwhile, the second heart field also consists of cardiac precursor cells present as separate cell populations on either side of the embryonic midline. They follow cues toward either the anterior (future head) or posterior (future tail) end of the previously formed heart tube to form ultimately other structures of the heart over the coming weeks, including left and right atria, right ventricle, coronary blood vessels, and segments of the great blood vessels of the heart.
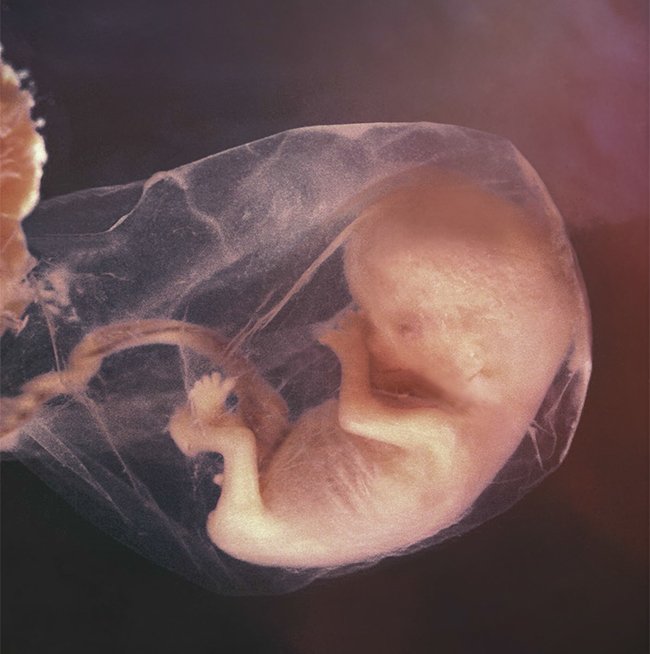
Photo © Life Issues Institute
Miracle Moments
The Placenta
Because the baby is a separate person who can’t share his mother’s blood, he receives nutrients through a temporary organ called the placenta. After the fertilized egg divides several times, the embryo will implant in the wall of the uterus, and the outer cell layer builds the placenta, whereas the other cells form the baby. Interestingly, both the baby’s cells and the mother’s cells build the placenta, so this life-sustaining organ functions by intricate coordination.
Though both the mother’s and baby’s blood flow through the placenta, they do not mix. Otherwise, the mother’s immune system would fight the foreign blood with antibodies. Instead, the complex arrangement of arteries and veins in the placenta allows the mother’s blood to exchange oxygen and nutrients with the baby.
While the baby’s organs develop, the placenta functions as the lungs, kidneys, digestive system, liver, and immune system.
After delivery, the placenta disconnects, severing vessels in the wall of the uterus. In God’s wise design, the muscles of the uterus tighten around torn blood vessels to control bleeding until the vessels can heal. This limits maternal blood loss.
How One Heart Tube Becomes Four Chambers
A baby’s rudimentary heart will start beating about three weeks after conception, around the time the mother begins to suspect she is pregnant. This early embryonic heart is a pulsating tube, not at all resembling the complex adult heart with its four chambers, valves, and attendant blood vessels. Around the third and fourth weeks of development, the heart tube undertakes a complicated maneuver called heart looping. At first, the straight tube lies along the embryo’s anterior-posterior (head-to-tail) axis. To understand the necessity of heart looping, picture the sequence of regional fates along the tube from anterior to posterior: outflow tract, future ventricles, and future atria (inflow). It’s kind of an upside-down heart, relative to the adult organ, with the posterior tube regions fated to become atria and the relatively anterior tube regions fated to become ventricles. Through highly coordinated interactions between mediating molecules and the internal “skeleton” of the heart tube cells and the extracellular matrix (external support of cells), the posterior atrial end of the heart tube curves into a tight U-shape, migrating toward the anterior ventricular end of the heart tube. Upon completion of this complex movement, the future atria are now closely apposed to the developing outflow tracts (future aorta and pulmonary arteries) with the future ventricles relatively posterior. Now in the correct relative positions, these structures will fuse and form the “right-side up” adult heart.
Doesn’t this seem overly complicated? Why not just start out with four populations of cells, increase the number of cells, and then hollow them out to form the chambers of the heart? (Something similar is known in spinal cord development, for example.) Why all this two-separate-heart-fields-migration-heart-looping rigmarole to make the heart? It seems like increasing the number of steps would increase the probability of something going awry. The fact is, scientists don’t know the reason for the complex process. Perhaps a wise and loving Creator wanted to reveal his attributes in every age, increasing the difficulty of believing that a functional, life-giving heart could result from an undirected evolutionary process of natural selection acting on random mutations over long periods of time.
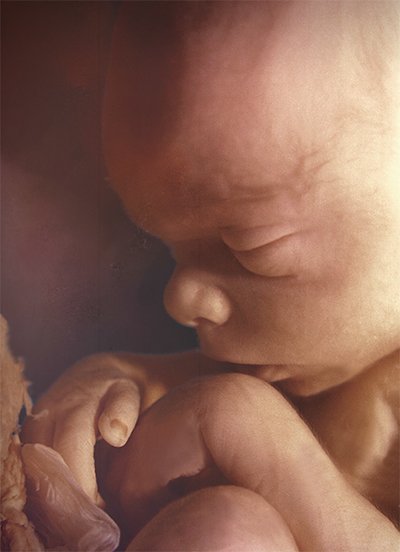
Photo © Life Issues Institute
Miracle Moments
Skin and Hair
At week 4, the baby begins some important transitions inside the womb, including gaining skin layers and hair. After all the skin layers develop, around week 15, the baby grows a hairline on its head. By birth, all hair follicles will be in place and perfectly spaced.
Until the final weeks of the pregnancy, the baby lacks body fat, so his skin appears transparent. Around 14 weeks, to help keep him warm, the baby’s body is covered in soft hair called lanugo. After the baby accumulates fat, the lanugo sheds around 30 weeks.
At 19 weeks, the baby’s fragile skin is coated in vernix, a white layer made of shed skin cells and oil, to protect him from the amniotic fluid. Closer to birth, this protective coating may decrease somewhat but continues to protect the baby’s skin until he reaches full term.
When the baby is ready to face the world, he is wrapped in warm layers of fat and hair.
The First Blood
The principle that life is in the blood has been known from ancient times, from Scriptures written by Moses under the inspiration of the Holy Spirit (Leviticus 17:11). Blood is life-giving and protective (immune system), and scientists nowadays have the technological tools to discover the fullness of this biological principle. In developmental biology, the origination of blood is one of the more fascinating stories. Blood cell development requires a process that is dependent on mechanical forces, specifically pressure and friction. During development, most processes are governed by cell communication, that is, molecules (e.g., proteins) interacting with each other and with the genome to cause a chain of events. However, a few processes, such as proper formation of the musculoskeletal systems, are driven by mechanotransduction (conversion of mechanical force into a molecular signal).
Here, the mechanical event of fluid flow activates a key developmental gene that leads to blood formation in the following way. The first blood stem cells are generated from the walls of the aorta, the largest blood vessel and outflow tract of the heart. The cells lining the aorta’s walls are called endothelial cells. As the embryonic heart pumps, the fluid flowing through the aorta (not yet blood but interstitial fluid, or “cell-bathing” fluid) applies shear forces (friction) to the endothelial cells. This friction force causes activation of the gene Runx1 exclusively in the endothelial cells of the ventral (belly) side of the aorta. Once activated, the Runx1 transcription factor converts these endothelial cells into blood stem cells that will migrate eventually to the liver and bone marrow. The absence of this critical factor prevents the production of all blood stem cells. So the next time you feel the press of friction forces in your life, remember blood cell development, and pray for patience and grace because God may be working something life-giving in you (see Romans 5:3–5).
Developmental Biology and the Origin of Life
Natural selection acting on random mutations over long periods of time would not sustain the embryo at each sequential step in the developmental process.
When God created Adam from the dust of the ground and Eve from Adam’s rib (Genesis 2:7, 21–22), he preloaded into our first parents’ genes the information for the process of developing new human beings. The examples from just this one organ system show that development is a sequentially dependent process. Each step of development is dependent upon the previous step. Disrupting any step in the process destroys the whole process so that the embryo will not reach reproductive adulthood. Therefore, it has zero “fitness” (relative number of offspring produced or differential reproduction), contributing no genes to the gene pool of the population. This sequential dependence of development shows that evolution fails as an explanatory mechanism for the origins of the vertebrate circulatory system, since natural selection acting on random mutations over long periods of time would not sustain the embryo at each sequential step in the developmental process. Instead, human beings—with their complex, integrated organ systems—were special creations of God.
The Ultimate Issue
God created mankind in his image. Therefore science—mankind—cannot measure the value of a human life. It is infinitely valuable to God. We know this because God paid an infinitely high price to redeem sinful humans. The price? The life of the eternal Son of God, Jesus Christ. God’s Word says, “Now in Christ Jesus you who once were far off have been brought near by the blood of Christ” (Ephesians 2:13). We see, then, human life is priceless because it can only be redeemed by the priceless blood of Christ.
Why is human life so valuable, causing God to spare not his Son but to give him up for us all while we were yet sinners (Romans 5:8, 8:32)? Because we were created in God’s image. Only mankind, of all God’s creation, was made in the image of God (Genesis 1:26). Therefore, to honor and to preserve human life is to honor God’s image; and to destroy human life is an egregious attack on the very image of God. This is why I take the position of opposing human embryo-destructive research, because it is illogical to destroy human life in order to save human life.
An important aspect of exploring the wonder of developmental biology is to foster this central truth of the sacredness of human life. In studying the scientific literature, students ask probing questions, such as, “Why don’t we know that?” I answer, “Because we don’t experiment on humans. Human beings have been created in the image of God, so we don’t do experiments that destroy human life.” Because of the strict limits placed on the research of human subjects, there are many things about human biology we do not know; and in order to revere the image of God in mankind, we must be content to allow that knowledge to remain cloaked in mystery.
However, experiments in developmental biology (e.g., adult cell culture studies and animal models) are very important for physicians and researchers. To diagnose certain syndromes, physicians must have excellent knowledge of the normal origin of tissues and organs so that a cluster of health problems may be uncovered by correctly linking them to their developmental root. Some cancer researchers are beginning to think about cancer as a “disease of development” because embryonic processes of cell proliferation and migration are dysregulated in adult tissues. Developmental genes being studied with great interest are those that promote normal migratory behavior in embryonic cells. Some of these genes, which are supposed to be “off” after development of the adult structure is completed, appear to be abnormally turned back “on” during the process of metastasis (i.e., when cancer cells leave the primary tumor to form a tumor elsewhere).
The development of a human or animal from a single cell is exceedingly complicated. Consider again the formation of the “double” embryonic heart, the complex process of heart-looping, and the special way in which blood cells are made. Each process involves dozens of factors that must be present in the right time and at the right place and active neither for too long nor too short a period. These events cry out, “Glory to God in the highest!” It is amazing that God thought of everything when he created. If he cares about every minute detail of weaving your physical body together, he also cares about every detail of your life—you, who are made in his image.
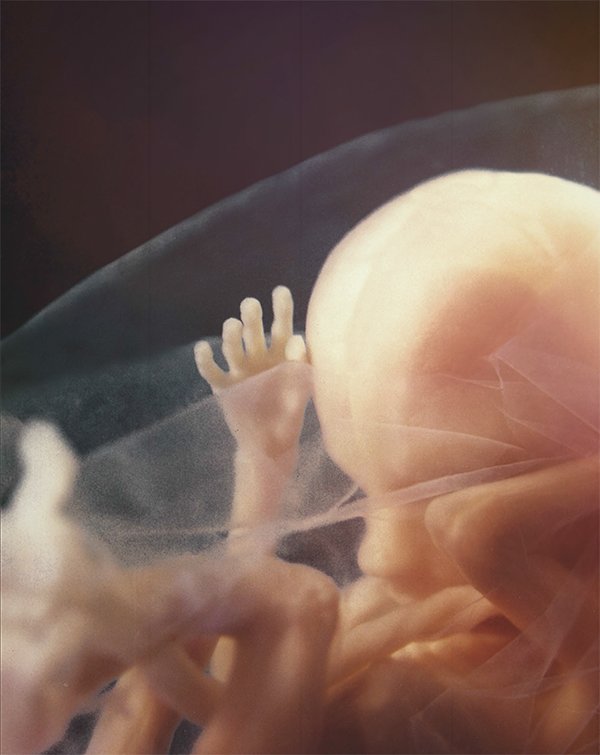
Photo © Life Issues Institute
Miracle Moments
From Liquid to Breath
For months, the placenta “breathes” for the baby. But at birth, a number of changes must occur in the lungs and circulatory system to allow the baby, who spent his life underwater, to breathe on his own.
For months, the lungs held fluid. Now the blood and lymph system remove the fluid and replace it with oxygen.
During development, blood bypassed the lungs, taking a shortcut through the heart where it must flow in order to be pumped to the rest of the body. At birth, the increase in oxygen causes the blood vessel that bypassed the lungs to close and sends blood flowing to the lungs. Also, the opening between right and left atria that was part of the bypass normally closes after birth.
Responding to the abrupt change in temperature and environment, the central nervous system prompts the lungs to take the first breath. All of this happens within seconds after birth.
has an earned doctorate in neurobiology and behavior with a minor in genetics and development from Cornell University and has held faculty positions at Grove City College, Pepperdine University, and Liberty University.
Answers Magazine
January–February 2019
An engineering feat that far surpasses landing a man on the moon is taking place all around us.
Browse Issue


Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis