

Tiktaalik photo by Eduard Solà, via Wikimedia Commons
The Fossil Record of ‘Early’ Tetrapods: Evidence of a Major Evolutionary Transition?
Originally published in Journal of Creation 17, no 2 (August 2003): 111-117.
Abstract
According to evolutionary theory, the origin of tetrapods from a fish-like ancestor during the Devonian Period was one of the major events in the history of life on earth.
Summary
According to evolutionary theory, the origin of tetrapods from a fish-like ancestor during the Devonian Period was one of the major events in the history of life on earth. The ‘drying pond’ hypothesis was proposed to explain the selection pressures behind the transition. According to this hypothesis, the tetrapods evolved as fishes became progressively better adapted to terrestrial conditions during prolonged episodes of drought. Recently, however, the assumption that feet and legs evolved to facilitate life on the land has been called into question. The ‘earliest’ known tetrapods with feet and legs are now thought to have been aquatic animals; evolutionists therefore argue that feet and legs evolved in a shallow water environment and were only later co-opted for use on the land. This paper reviews the radical changes in thinking about the fish-tetrapod transition that have taken place in the evolutionary community. It also considers the chimeromorphic nature of Devonian tetrapods and fishes, and offers some critical comments on the evolutionary interpretation of their fossil record.
Evolutionists believe that tetrapods—i.e. vertebrates with four limbs—were the first animals to move on to the land, having evolved from a fish ancestor during the Devonian period (conventionally 408 to 360 million years ago). The fossil record of Devonian tetrapods is often presented as compelling evidence of this major evolutionary transition.1 Science writer Carl Zimmer has written a popular book, At the Water’s Edge,2 which purports to show how life came ashore (i.e. how fish evolved into tetrapods) and then went back to the sea (i.e. how land mammals gave rise to the whales). A more technical presentation was written recently by Jenny Clack, Reader in Vertebrate Palaeontology and Senior Assistant Curator of the University Museum of Zoology, Cambridge. Entitled Gaining Ground: The Origin and Evolution of Tetrapods,3 it begins with these words:
‘About 370 million years ago, something strange and significant happened on Earth. That time, part of an interval of Earth’s history called the Devonian Period by scientists such as geologists and paleontologists, is known popularly as the Age of Fishes. After about 200 million years of earlier evolution, the vertebrates—animals with backbones—had produced an explosion of fishlike animals that lived in the lakes, rivers, lagoons, and estuaries of the time. The strange thing that happened during the later parts of the Devonian period is that some of these fishlike animals evolved limbs with digits—fingers and toes. Over the ensuing 350 million years or so, these so-called tetrapods gradually evolved from their aquatic ancestry into walking terrestrial vertebrates, and these have dominated the land since their own explosive radiation allowed them to colonize and exploit the land and its opportunities. The tetrapods, with their limbs, fingers, and toes, include humans, so this distant Devonian event is profoundly significant for humans as well as for the planet.’4
Indeed, according to the cladistic framework that now dominates evolutionary systematics, humans are not simply descended from fish—they are fish! Clack states:
‘Although humans do not usually think of themselves as fishes, they nonetheless share several fundamental characters that unite them inextricably with their relatives among the fishes … Tetrapods did not evolve from sarcopterygians [lobe-finned fishes]; they are sarcopterygians, just as one would not say that humans evolved from mammals; they are mammals.’5
In this paper I will critically examine the fossil record of ‘early’ tetrapods and discuss the way in which older evolutionary views of their origin have been overturned in the last two decades. I will also consider the mosaic distribution of characters that we observe in Devonian tetrapods and fishes, the problems that it poses for evolutionary theory, and how it might be understood in a creationist framework.
The ‘drying pond’ hypothesis
Many evolutionary scenarios have been proposed to explain the origin of tetrapods. Most of them were developed to answer the question, ‘Why did fish leave the water and come onto the land?’ The early theories usually focused on the environmental setting and selection pressures behind the transition. Tetrapods were thought to have evolved during the Devonian, a period associated in many parts of the world with sediments stained red by iron oxide. Classic red beds, such as the Siluro-Devonian rocks of Europe (the Old Red Sandstone) and their North American equivalents (the Catskill and Escuminac formations), have often been interpreted as the product of hot, semi-desert environments with seasonal wetness. This led many to speculate that an increasingly arid climate was a major influence on the evolution of air-breathing vertebrates. A classic paper by Barrell6 set the scene for much future discussion. He argued that the first tetrapods arose ‘under the compulsion of seasonal dryness’.7 Under such conditions, it was suggested, the air-bladder of certain fishes became progressively better adapted as an organ of respiration and the gills atrophied. The development of a new system of breathing allowed fishes to survive the drought conditions by moving between bodies of water. Those fishes with more limb-like appendages were better able to make the journey and this ultimately led to the evolution of limbs with digits. This became known as ‘the drying pond hypothesis’ and was popularized by the great vertebrate palaeontologist Alfred Sherwood Romer.8
‘Early’ tetrapods from East Greenland
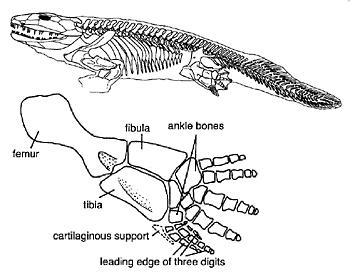
Figure 1. Reconstruction of Ichthyostega, showing skull, vertebral column, and limbs, and its hind limb based on a specimen collected in 1987. Note the seven digits on the hind limb (from Clack).15
When Romer was popularizing the ‘drying pond’ idea, the earliest known tetrapods were Ichthyostega and Acanthostega from the Upper Devonian of East Greenland. Ichthyostega was first described by Säve-Söderbergh9 and then by Jarvik in a series of papers and a monograph.10–12 Although the anatomy of Ichthyostega is known in considerable detail, its body proportions are uncertain because the fossil material comes from more than one individual. Ichthyostega is about one metre long with a broad, flat head, short, barrel-shaped body, stocky legs, large pelvic and pectoral girdles, and a rib cage with broad, overlapping ribs (Figure 1). It is very evidently a tetrapod, with limbs rather than fins. Nevertheless, Ichthyostega has some fish-like characteristics, including a lateral line system and a tail with bony fin rays. Early reconstructions portrayed Ichthyostega as a semi-aquatic creature but most later ones depicted it as a predominantly terrestrial animal (e.g. Jarvik13). As recently as 1988, a major vertebrate palaeontology text described Ichthyostega as a fairly typical land animal with the usual complement of five digits on the hind limb.14 The second Devonian tetrapod from East Greenland was Acanthostega.9,10 For many years this animal was known only from two partial skull roofs, but these were enough to mark it out as different from Ichthyostega.
The search for evolutionary ancestors
Evolutionists sought the ancestry of the tetrapods among the lobe-finned fishes. Although the lobe-fins are dominant in the fossil fish faunas of the Palaeozoic (conventionally 590 to 248 million years ago), they are represented today by only four surviving genera (the coelacanth Latimeria and three genera of lungfish). In 1892, Cope and others argued that tetrapods had evolved from the crossopterygians, the group of lobe-fins that includes the coelacanths.16 Various crossopterygians were proposed as the ‘model ancestor’, including Sauripteris17,18 and Osteolepis.19 However, most attention settled upon Eusthenopteron, from Escuminac Bay in Quebec, Canada. This is the fish that was commonly illustrated, in popular books on fossils, as hauling itself up onto Devonian riverbanks (e.g. Owen20).
Nevertheless, there was evidently a substantial discontinuity in the fossil record between terrestrial vertebrates like Ichthyostega and their presumed ancestors. This was reflected in creationist treatments of the problem21 and acknowledged by evolutionists, such as Carroll22 who wrote:
‘We have not found any fossils that are intermediate between such clearly terrestrial animals and the strictly aquatic rhipidistians described in the previous chapter.’
| Taxon | Stratigraphic unit | Age | Location | Material | Reference(s) |
|---|---|---|---|---|---|
| Pederpes | Ballagan Fm | Tournaisian | Scotland | Skull, almost complete articulated skeleton | 23 |
| Sinostega | Zhongning Fm | Famennian | Ningxia Hui, China | Incomplete left mandible | 24 |
| Tulerpeton | Khovanshchina Beds | Famennian | Tula Region, Russia | Fore and hind limbs, partial pectoral and pelvic girdles, skull fragments | 25–28 |
| Ventastega | Ketleri Fm | Famennian | Latvia | Skull fragments, girdle fragments | 29 |
| Acanthostega | Britta Dal Fm | Famennian | East Greenland | Skulls, articulated skeletons | 9,10,30–36,44,50 |
| Ichthyostega | Aina Dal Fm Britta Dal Fm | Famennian | East Greenland | Skulls, skeletal elements, some articulated | 9–12,44 |
| Hynerpeton | Catskill Fm | Famennian | Pennsylvania, USA | Pectoral girdle, skull fragments | 37,38 |
| Densignathus | Catskill Fm | Famennian | Pennsylvania, USA | Lower jaw | 38 |
| Metaxygnathus | Cloghnan Shale | Famennian | New South Wales, Australia | Lower jaw | 39 |
| Elginerpeton | Scat Craig Beds | Frasnian | Scotland | Ilia, limb bones, skull and pectoral girdle fragments | 40–42 |
| Obruchevichthys | Ogre Beds | Frasnian | Latvia | Lower jaw fragments | 40 |
| Livoniana | Gauja Fm | Givetian | Latvia | Lower jaw fragments | 43 |
Aquatic tetrapods challenge the ‘drying pond’ hypothesis
Since 1990 our knowledge of ‘early’ tetrapods has been greatly expanded, with many new taxa being described. Fossil material is now known from Scotland, Greenland, Latvia, the USA, Australia, Russia, and China (Table 1).23–43 Furthermore, our understanding of the Greenland tetrapods has been revolutionized by the discovery of new material. As a consequence, a major re-evaluation of tetrapod origins has taken place, and almost every aspect of the ‘drying pond’ hypothesis has had to be discarded.
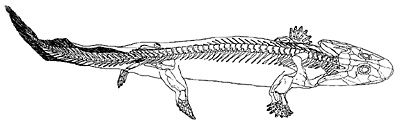
Figure 2. Acanthostega in a swimming posture (from Clack).46
The fatal blow to the ‘drying pond’ hypothesis has been the realization that the Devonian tetrapods were predominantly aquatic in habit. New ichthyostegid material, including a well-preserved and articulated hind limb, collected by an expedition to East Greenland in 1987, revealed that Ichthyostega was polydactylous, with seven digits on the hind limb (Figure 1).44 This was a very surprising discovery because pentadactyly had been assumed to be the normal condition in ‘early’ tetrapods. Furthermore, the flattened bones and inflexible ankle of the hind limb suggests that it was more like the paddle of an elephant seal than the leg of a terrestrial animal.45 It appears that the earliest reconstruction of Ichthyostega as a creature at home in the water was more accurate than later ones portraying it on land.
Acanthostega is also much more completely known as a result of material collected by the 1987 expedition, including the first postcranial remains.47,48 It was a smaller animal than Ichthyostega and its teeth suggest that it had a different diet. Several articulated specimens were found in a single lens of rock, interpreted as a possible flash flood deposit.49 The remarkable preservation meant that some delicate structures, not often preserved in fossil tetrapods, are known in Acanthostega. The gill skeleton was fish-like50 and it has been suggested that Acanthostega had internal gills somewhat similar to those of the Australian lungfish (Neoceratodus). Acanthostega had a tail with fin rays, even larger than that of Ichthyostega (Figure 2). The fin rays also extended further beneath the tail, in similar fashion to those of a lungfish, suggesting that Acanthostega was a thoroughly aquatic creature. This conclusion is supported by the morphology of the fore and hind limbs which are difficult to interpret as load-bearing structures; rather, they appear to be designed for swimming. As with Ichthyostega, perhaps the most extraordinary feature was the number of digits. An articulated fore limb revealed eight digits in a paddle-like arrangement (Figure 3). Clack51 speculates that they may have been enclosed in some kind of webbing.
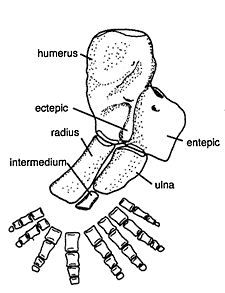
Figure 3. The left forelimb of Acanthostega, showing the eight digits (from Clack).52
Most evolutionists had assumed that the origin of limbs with digits was synonymous with the vertebrate invasion of the land. This led to the popular ‘conquest of the land’ idea, typified by artistic reconstructions and museum displays of fish crawling out of Devonian pools. However, the latest thinking about the aquatic or semi-aquatic nature of the Devonian tetrapods has led modern-day evolutionists to reject this assumption. They now argue that the key tetrapod characters evolved for a shallow-water existence and were only later co-opted for terrestrial use. The new generation of Darwinists dismisses the ‘drying pond’ hypothesis as untestable story-telling, and increasingly relies on cladistics as an alternative framework for understanding the transition. The cladistic approach to the fish-tetrapod transition focuses on determining the sequence of acquisition of key tetrapod characteristics, from which inferences are drawn about the nature of the transition.53 We should recognize, however, that the cladistic methodology is inherently Darwinian and assumes from the outset the continuity of life. By its very nature, cladistics is insensitive to the discontinuities which creationists believe characterize living things.54
Other problems with the ‘drying pond’ hypothesis
The drying pond hypothesis has other problems.55 For instance, it is recognized that red beds are not necessarily indicators of arid climates:
‘The red bed problem has been extremely controversial, with marked differences of opinion, possibly due to the fact that the term “red bed” is a catchall for many sedimentary types produced under different conditions, the only common feature of them being the red color.’56
Modern red beds develop in the oxidizing conditions of the low latitude tropics (e.g. the Amazon Basin). Such environments are characterized by monsoonal rainfall, not arid conditions. Another problem is that, even if the red beds were laid down under conditions of semi-aridity, evolutionists cannot assume that the tetrapods arose in such environments, for the simple reason that many Devonian sediments are not red beds. Some are interpreted as river, lake, or near-shore sediments rich in organic matter, suggesting nearby forests.57
Furthermore, a survey of modern fishes that leave the water to spend time on land58 affords no support for the ‘drying pond’ hypothesis. There is no association between those that leave the water and those that possess digit-like fins. For example, eels undertake long journeys overland but they have nothing that could be described as digit-like appendages. Indeed, most of the fishes that possess digit-like structures are deep water species or habitual bottom dwellers, such as the Sargassum frogfish.
New views on tetrapod ancestry
There have also been changes of opinion about which group of fishes is closest to the ancestry of tetrapods. Eusthenopteron is no longer regarded as the model ancestor. Depictions showing this fish emerging onto dry land owed more to evolutionary presuppositions than evidence. Eusthenopteron was a rather undistinguished fish with no obvious adaptations to terrestrial life; tetrapod-like behaviour was attributed to it simply because there was no better candidate to fill the role of tetrapod ancestor. The true lifestyle of Eusthenopteron seems to have been that of a lurking aquatic predator, somewhat similar to the modern pike (Esox).
Attention is now focused on the formerly more obscure lobe-finned fishes, Panderichthys and Elpistostege. Until recently, these two genera were united in a family called the panderichthyids, but evolutionists now believe that they are not uniquely related to each other.59 Fossil material from Latvia and Canada shows that these fish were more tetrapod-like than other lobe-fins. Indeed, based on a partial skull roof, Elpistostege was originally described as a tetrapod.60 Although there has been dissent,61,62 these genera are increasingly regarded by evolutionists as the closest known relatives of tetrapods.63–65 The latest work by Ahlberg et al.43 indicates that Elpistostege is even more tetrapod-like than Panderichthys. These fish have crocodile-like skulls with dorsally placed eyes, straight tails, and slightly flattened bodies without dorsal or anal fins (see Figure 4). Like tetrapods, but unlike all other fishes, they also have frontal bones in the skull roof. Like Eusthenopteron, they seem designed for life as shallow-water predators.

Figure 4. Panderichthys, an Upper Devonian lobe-finned fish regarded by evolutionists as close to the ancestor of tetrapods (from Clack).59
Chimeromorphs pose problems for evolutionary theory
Creationists and evolutionists have observed that many organisms, both fossil and living, exhibit a mosaic distribution of character traits. Parker66 put it this way:
‘Each created kind is a unique combination of traits that are individually shared with members of other groups.’
Stephen Jay Gould called such organisms ‘mosaic forms’ or ‘chimeras’67 while Kurt Wise68,69 calls them chimeromorphs. The duck-billed platypus (Ornithorhynchus anatinus), for instance, has features of both mammals (hair, milk production) and reptiles (egg-laying). Perhaps the best-known fossil example is Archaeopteryx, which combines feathers with teeth and wing claws. In fact, a mosaic pattern of character distribution is seen in many other fossil organisms. For instance, Woodmorappe70 recently drew attention to the chimeric nature of the pakicetids, a group of terrestrial artiodactyls with a whale-like inner ear.
This observation seems to apply to the Devonian tetrapods and fishes considered in this article. For example, Daeschler et al. noted that:
‘Devonian tetrapods show a mosaic of terrestrial and aquatic adaptations.’71
Some of the fishes possess tetrapod-like characters while the tetrapods have fish-like features. Evolutionists interpret mosaic organisms like these as evolutionary intermediates linking major groups. However, Wise72 makes an important point against this interpretation:
‘Although the entire organism is intermediate in structure, it’s the combination of structures that is intermediate, not the nature of the structures themselves. Each of these organisms appears to be a fully functional organism full of fully functional structures.’
Evolutionary theory might lead us to expect examples of intermediate structures, but there is nothing intermediate about, for example, the internal gills of Acanthostega, its lateral line system, or its limbs. They are fully developed and highly complex. What is unusual is their combination in a single organism. Intelligent design offers an alternative understanding of this widespread pattern. The Devonian tetrapods are thought to have lived a predatory lifestyle in weed-infested shallow water. They were therefore equipped with characteristics appropriate to that habitat (e.g. crocodile-like morphology with dorsally placed eyes, limbs and tails made for swimming, internal gills, lateral line systems). Some of these features are also found in fishes that shared their environment.
The mosaic pattern makes it difficult to identify organisms or groups of organisms that possess the ‘right’ combination of characters to be considered part of an evolutionary lineage. Consider the tetrapod-like lobe-fins Panderichthys and Elpistostege. Despite their appearance, these fish have some unique characters (such as the design of the vertebrae) that rule them out as tetrapod ancestors. At best, evolutionists can only claim that they are a model of the kind of fish that must have served as that ancestor. The same problem is encountered with the Devonian tetrapods. For example, Ichthyostega is described as ‘a very strange animal, and parts of it are like no other known tetrapod or fish’.73 Similarly, the shoulder girdles of the Devonian tetrapods ‘are not obviously halfway in structure between those of fishes and those of later tetrapods but have some unique and some unexpected features’.74 Another example is Livoniana, a so-called ‘near tetrapod’ known from two lower jaw fragments. It possesses a curious mixture of fish-like and tetrapod-like characteristics, but it also has up to five rows of teeth, a feature not seen either in the fishes from which it is thought to be descended nor the tetrapods into which it is said to be evolving.75 That the mosaic distribution of characters can cause great confusion is exemplified by the recent discovery of Psarolepis, a fish from the Upper Silurian/Lower Devonian of China, which combines characters found in placoderms, chondrichthyans, ray finned fishes, and lobe-fins.76
Additional problems with ‘early’ tetrapod evolution
Another problem is that the fossil record imposes tight constraints on the timing of the supposed transition. The earliest tetrapod fossils are found in late Frasnian sediments, but their presumed ancestors are hardly much older. To exacerbate the situation, the Frasnian ‘near tetrapods’ (Obruchevichthys, Elginerpeton, Livoniana) are already morphologically diverse at their first appearance.77 Thus Darwinists are compelled to postulate a rapid burst of evolution in which radical changes must have taken place:
‘Panderichthys and Elpistostege flourished in the early Frasnian and are some of the nearest relatives of tetrapods. But tetrapods appear only about 5 to 10 million years later in the late Frasnian, by which time they were widely distributed and had evolved into several groups, including the lineage leading to the tetrapods of the Famennian. This suggests that the transition from fish to tetrapod occurred rapidly within this restricted time span.’78
Second, key morphological transitions, such as the purported change from paired fins to limbs with digits, remain undocumented by fossils. Where appendages are known they are clearly either fish-like fins or digit-bearing limbs, not at some transitional stage from one to the other. At one time it was claimed that the pectoral fins of rhizodonts, a group of lobe-finned fish, were remarkably similar to tetrapod limbs, but following the description of Gooloogongia from the Famennian of New South Wales, Johanson and Ahlberg79 have urged that they not be used as a model for the origin of tetrapod limbs. Furthermore, the pectoral fins of lobe-finned fish tend to be larger than the pelvic fins, whereas the Devonian tetrapods were ‘rear-wheel drive’ animals with larger hind limbs than fore limbs.80 None of the recent fossil discoveries shed any light on this supposed reconfiguration.
Third, there are functional challenges to Darwinian interpretations. For instance, in fish the head, shoulder girdle, and circulatory systems constitute a single mechanical unit. The shoulder girdle is firmly connected to the vertebral column and is an anchor for the muscles involved in lateral undulation of the body, mouth opening, heart contractions, and timing of the blood circulation through the gills.81 However, in amphibians the head is not connected to the shoulder girdle, in order to allow effective terrestrial feeding and locomotion. Evolutionists must suppose that the head became incrementally detached from the shoulder girdle, in a step-wise fashion, with functional intermediates at every stage. However, a satisfactory account of how this might have happened has never been given.
Conclusion
Recent discoveries have undoubtedly advanced our knowledge of Devonian tetrapods and future creationist discussions of tetrapod origins must take this into account. It is no longer sufficient for creationists to contrast Eusthenopteron with Ichthyostega and point to the large morphological gap between them. We need to have more to say. Nevertheless, the presumed transition from fish to tetrapods remains contentious. The data and their interpretation are a source of lively debate and ongoing controversy:
‘In the not-too-distant past, there was almost no fossil material, and ideas were based largely on informed guesswork. Speculation was intense, and as is often the case, in inverse proportion to the amount of data. To be truthful, there is still not much real data, so that speculation is still active, and whatever is concluded today may be overturned by the discovery of a new fossil tomorrow. That in some sense is to be hoped for, because only in that way can guesses be falsified and tested as scientific hypotheses.’82
A robust rationale for concluding that the Upper Devonian tetrapods evolved from a fish ancestor, or that they gave rise to Carboniferous tetrapod lineages, is lacking. It is hoped that this paper may stimulate creationists to develop a fuller understanding of these remarkable creatures and their ecological and geological context.83
About the Author
Paul Garner has a B.Sc. (Hons) in Geology and Biology and is a Fellow of the Geological Society of London. He works full-time as a speaker and researcher with Biblical Creation Ministries in the UK. He is also a Committee Member of the Biblical Creation Society, co-editor of the BCS journal, Origins, and is on the Board of The Genesis Agendum, a charitable company promoting church and public awareness of the substantial historical and scientific evidence supporting the biblical record.
References
- Morton, G.R., Fish to amphibian transition, <home.entouch.net/dmd/transit.htm>, 27 June 2003. Return to text.
- Zimmer, C., At the Water’s Edge: Fish with Fingers, Whales with Legs, and How Life Came Ashore but Then Went Back to Sea, Touchstone, New York, 1998. Return to text.
- Clack, J.A., Gaining Ground: The Origin and Evolution of Tetrapods, Indiana University Press, Bloomington, 2002. Return to text.
- Clack, Ref. 3, p. 1. Return to text.
- Clack, Ref. 3, pp. 17, 18. Return to text.
- Barrell, J., Influence of Silurian-Devonian climates on the rise of air-breathing vertebrates, Bulletin of the Geological Society of America 27:387–436, 1916. Return to text.
- Barrell, Ref. 6, p. 390. Return to text.
- Romer, A.S., Man and the Vertebrates, Volume 1, Penguin, Harmondsworth, pp. 48–49, 1954. Return to text.
- Säve-Söderbergh, G., Preliminary note on Devonian stegocephalians from East Greenland, Meddelelser om Grønland 94:1–107, 1932. Return to text.
- Jarvik, E., On the fish-like tail in the ichthyostegid stegocephalians, Meddelelser om Grønland 114:1–90, 1952. Return to text.
- Jarvik, E., Specializations in early vertebrates, Annales Societe Royale Zoologique de Belgique 94:11–95, 1965. Return to text.
- Jarvik, E., The Devonian tetrapod Ichthyostega, Fossils and Strata 40:1–213, 1996. Return to text.
- Jarvik, E., Basic Structure and Evolution of Vertebrates, Volumes 1 and 2, Academic Press, New York, 1980. Return to text.
- Carroll, R.L., Vertebrate Paleontology and Evolution, W.H. Freeman and Company, New York, p. 164, 1988. Return to text.
- Clack, Ref. 3, p. 114. Return to text.
- Cope, E.D., On the phylogeny of the vertebrata, Proceedings of the American Philosophical Society 30:278–281, 1892. Return to text.
- Gregory, W.K., Present status of the problem of the origin of the Tetrapoda, with special reference to the skull and paired limbs, Annals of the New York Academy of Sciences 26:317–383, 1915. Return to text.
- Gregory, W.K., Further observations on the pectoral girdle and fin of Sauripterus taylori Hall, a crossopterygian fish from the Upper Devonian of Pennsylvania, with special reference to the origin of the pentadactylate extremities of Tetrapoda, Proceedings of the American Philosophical Society 75:673–690, 1935. Return to text.
- Watson, D.M.S., Croonian lecture—The evolution and origin of the Amphibia, Philosophical Transactions of the Royal Society of London Series B 214:189–257, 1926. Return to text.
- Owen, E., Prehistoric Animals: the Extraordinary Story of Life Before Man, Octopus Books, London, p. 20, 1975. Return to text.
- Gish, D.T., Evolution: The Fossils Say No! Third edition, Creation-Life Publishers, San Diego, pp. 78–83, 1979. Return to text.
- Carroll, Ref. 14, p. 158. Return to text.
- Clack, J.A., An early tetrapod from ‘Romer’s Gap’, Nature 418:72–76, 2002. Return to text.
- Zhu, M., Ahlberg, P.E., Zhao, W. and Jia, L., First Devonian tetrapod from Asia, Nature 420:760–761, 2002. Return to text.
- Lebedev, O.A., The first record of a Devonian tetrapod in the USSR, Doklady Akademii Nauk SSSR 278:1470–1473, 1984. Return to text.
- Lebedev, O.A., The first tetrapods: searchings and findings, Priroda 11:26–36, 1985. Return to text.
- Lebedev, O.A. and Clack, J.A., Upper Devonian tetrapods from Andreyevka, Tula region, Russia, Palaeontology 36:721–734, 1993. Return to text.
- Lebedev, O.A. and Coates, M.I., The postcranial skeleton of the Devonian tetrapod Tulerpeton curtum Lebedev, Zoological Journal of the Linnean Society 114:307–348, 1995. Return to text.
- Ahlberg, P.E., Lukševis, E. and Lebedev, O., The first tetrapod finds from the Devonian (Upper Famennian) of Latvia, Philosophical Transactions of the Royal Society of London Series B 343:303–328, 1994. Return to text.
- Clack, J.A., New material of the early tetrapod Acanthostega from the Upper Devonian of East Greenland, Paleontology 31:699–724, 1988. Return to text.
- Clack, J.A., Discovery of the earliest-known tetrapod stapes, Nature 342:424–427, 1989. Return to text.
- Clack, J.A., Acanthostega gunnari, a Devonian tetrapod from Greenland; the snout, palate and ventral parts of the braincase, with a discussion of their significance, Meddelelser om Grønland: Geoscience 31:1–24, 1994. Return to text.
- Clack, J.A., Earliest known tetrapod braincase and the evolution of the stapes and fenestra ovalis, Nature 369:392–394, 1994. Return to text.
- Coates, M.I., The Devonian tetrapod Acanthostega gunnari Jarvik: postcranial anatomy, basal tetrapod interrelationships and patterns of skeletal evolution, Transactions of the Royal Society of Edinburgh: Earth Sciences 87:363–421, 1996. Return to text.
- Ahlberg, P.E. and Clack, J.A., Lower jaws, lower tetrapods—a review based on the Devonian genus Acanthostega, Transactions of the Royal Society of Edinburgh: Earth Sciences 89:11–46, 1998. Return to text.
- Clack, J.A., The neurocranium of Acanthostega gunnari Jarvik and the evolution of the otic region in tetrapods, Zoological Journal of the Linnean Society 122:61–97, 1998. Return to text.
- Daeschler, E.B., Shubin, N.H., Thomson, K.S. and Amaral, W.W., A Devonian tetrapod from North America, Science 265:639–642, 1994. Return to text.
- Daeschler, E.B., Early tetrapod jaws from the Late Devonian of Pennsylvania, Journal of Paleontology 74:301–308, 2000. Return to text.
- Campbell, K.S.W. and Bell, M.W., A primitive amphibian from the Late Devonian of New South Wales, Alcheringa 1:369–381, 1977. Return to text.
- Ahlberg, P.E., Tetrapod or near-tetrapod fossils from the Upper Devonian of Scotland, Nature 354:298–301, 1991. Return to text.
- Ahlberg, P.E., Elginerpeton pancheni and the earliest tetrapod clade, Nature 373:420–425, 1995. Return to text.
- Ahlberg, P.E., Postcranial stem tetrapod remains from the Devonian of Scat Craig, Morayshire, Scotland, Zoological Journal of the Linnean Society 122:99–141, 1998. Return to text.
- Ahlberg P.E., Lukševičs, E. and Mark-Kurik, E., A near-tetrapod from the Baltic Middle Devonian, Palaeontology 43:533–548, 2000. Return to text.
- Coates, M.I. and Clack, J.A., Polydactyly in the earliest known tetrapod limbs, Nature 347:66–69, 1990. Return to text.
- Clack, Ref. 3, p. 119. Return to text.
- Clack, Ref. 3, p. 121. Return to text.
- Clack, J.A., Pioneers of the land in East Greenland, Geology Today 4:407–409, 1988. Return to text.
- Bendix-Almgreen, S.E., Clack, J.A. and Olsen, H., Upper Devonian tetrapod palaeoecology in the light of new discoveries in East Greenland, Terra Nova 2:131–137, 1990. Return to text.
- Clack, Ref. 3, p. 110. Return to text.
- Coates, M.I. and Clack, J.A., Fish-like gills and breathing in the earliest known tetrapod, Nature 352:234–236, 1991. Return to text.
- Clack, Ref. 3, p. 126. Return to text.
- Clack, Ref. 3, p. 125. Return to text.
- Ahlberg, P.E. and Milner, A.R., The origin and early diversification of tetrapods, Nature 368:507–514, 1994; p. 507. Return to text.
- Wise, K.P., Baraminology: a young-Earth creation biosystematic method; in: Walsh, R.E. and Brooks, C.L. (Eds), Proceedings of the Second International Conference on Creationism, Volume II, Creation Science Fellowship, Pittsburgh, pp. 345–360, 1990; p. 346. Return to text.
- Clack, Ref. 3, pp. 99–104. Return to text.
- Krynine, P.D., The origin of red beds, Transactions of the New York Academy of Sciences 11:60–68, 1949; p. 60. Return to text.
- Clack, Ref. 3, p. 101. Return to text.
- Sayer, M.D.J. and Davenport, J., Amphibious fish: why do they leave water? Reviews in Fish Biology and Fisheries 1:159–181, 1991. Return to text.
- Clack, Ref. 3, p. 62. Return to text.
- Westoll, T.S., Ancestry of the tetrapods, Nature 141:127–128, 1938. Return to text.
- Chang, M-M., ‘Rhipidistians’, dipnoans, and tetrapods; in: Schultze, H-P. and Trueb L. (Eds), Origins of the Higher Groups of Tetrapods: Controversy and Consensus, Comstock Publishing Associates, Ithaca, New York, pp. 3–28, 1991. Return to text.
- Panchen, A.L. and Smithson, T.R., Character diagnosis, fossils, and the origin of tetrapods, Biological Reviews 62:341–438, 1987. Return to text.
- Schultze, H-P. and Arsenault, M., The panderichthyid fish Elpistostege: a close relative of tetrapods? Palaeontology 28:293–309, 1985. Return to text.
- Vorobyeva, E.I., Morphology and nature of evolution of crossopterygian fishes, Trudy Paleontological Institute Akademie Nauk SSSR 163:1–239, 1977. Return to text.
- Vorobyeva, E. and Schultze, H-P., Description and systematics of panderichthyid fishes with comments on their relationship to tetrapods; in: Schultze, H-P. and Trueb, L. (Eds), Origins of the Higher Groups of Tetrapods: Controversy and Consensus, Comstock Publishing Associates, Ithaca, New York, pp. 68–109, 1991. Return to text.
- Parker, G., Creation: Facts of Life, Master Books, Green Forest, p. 118, 1994. Return to text.
- Wise, K.P., The origin of life’s major groups; in: Moreland, J.P. (Ed.), The Creation Hypothesis: Scientific Evidence for an Intelligent Designer, InterVarsity Press, Downers Grove, pp. 211–234, 1994; p. 227. Return to text.
- Wise, K.P., Towards a creationist understanding of transitional forms, TJ 9(2):216–222, 1995. Return to text.
- Wise, K.P., Is life singularly nested or not? in: Walsh, R.E. (Ed.), Proceedings of the Fourth International Conference on Creationism, Creation Science Fellowship, Pittsburgh, pp. 619–631, 1998. Return to text.
- Woodmorappe, J., Walking whales, nested hierarchies, and chimeras: do they exist? TJ 16(1):111–119, 2002. Return to text.
- Daeschler et al., Ref. 37, p. 641. Return to text.
- Wise, Ref. 67, p. 227. Return to text.
- Clack, Ref. 3, p. 115. Return to text.
- Clack, Ref. 3, p. 42. Return to text.
- Lamb, A., Livoniana—have they (finally!) found a missing link? TJ 16(1):4–6, 2002. Return to text.
- Zhu, M., Yu, X. and Janvier, P., A primitive fossil fish sheds light on the origin of bony fishes, Nature 397:607–610, 1999. Return to text.
- Clack, Ref. 3, p. 137. Return to text.
- Clack, Ref. 3, p. 96. Return to text.
- Johanson, Z. and Ahlberg, P.E., A complete primitive rhizodont from Australia, Nature 394:569–573, 1998. Return to text.
- Ahlberg and Milner, Ref. 53, p. 509. Return to text.
- Gudo, M. and Homberger, D.G., Functional morphology of the coracoid bar of the Spiny Dogfish (Squalas acanthias): implications for the evolutionary history of the shoulder girdle of vertebrates, 43rd Annual Meeting of the Palaeontological Association, Manchester, 19–22 December 1999; <www.palass.org/index.html>, 4 April 2003. Click on ‘Abstracts’ then ‘Manchester 1999’. Return to text.
- Clack, Ref. 3, p. 3. Return to text.
- One approach to understanding the ‘early’ tetrapods is outlined in a forthcoming paper: Garner, P., From Fins to Feet: Did Fish Evolve into Tetrapods? The Genesis Agendum Occasional Paper 9, 2003. Available from The Genesis Agendum, P.O.Box 5918, Leicester LE2 3XE, UK. Return to text.
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis