

Chapter 14
Fossil Record Evolution: Any Transitional Forms?
Is there such a thing as fossil record evolution? The central idea of evolution is that all of the kinds of living organisms on earth share a common ancestor and that over time they have evolved one from another by an unplanned and unguided natural process. This unobserved sort of “amoeba-to-man” evolution extending over hundreds of millions of years is called macroevolution to distinguish it from the relatively small-scale variations we observe among the individuals of a species. Evolutionists like to refer to these small variations as “microevolution” with the tacit assumption that over eons of time they add up incrementally to produce macroevolution. Thus, evolutionists look for evidence of these incremental steps, often referring to them as “transitional forms,” suggesting that they represent stages of transformation of one organism into a different kind of organism.
Since macroevolution is not observable in the time frame of human observers, evolutionists often invoke microevolution as both evidence for macroevolution as well as its presumed mechanism. But as any animal or plant breeder knows, the limited variation that is observed among the individuals of a species has not been observed to lead to the essentially limitless process of macroevolution. In 1980, a group of evolutionists met in Chicago to discuss the relationship of micro- and macroevolution. Roger Lewin summed up this meeting in the journal Science as follows:
The central question of the Chicago conference was whether the mechanisms underlying microevolution can be extrapolated to explain the phenomena of macroevolution. At the risk of doing violence to the positions of some of the people at the meeting, the answer can be given as a clear No.1
The lack of a clear relationship between microevolution and macroevolution has continued to be a problem for evolutionists.2
No matter what mechanism one might postulate for macroevolution, in the course of presumed evolutionary history there would have been an unimaginably vast number of transitional forms revealing at least some of the incremental stages of macroevolution. Thus evolutionists typically turn to the fossil record in an effort to identify transitional stages in the macro evolutionary process. When this fails, they turn to currently living biological organisms in the hope of “reconstructing” evolutionary transitional stages from living examples. When an appearance of progress is lacking among living organisms and their organs, evolutionists turn to artists who obligingly illustrate what they believe must surely have been the missing transitional stages of evolutionary progress. And, finally, when even artistic imagination fails to produce plausible intermediates of evolutionary progress, some evolutionists simply deny that there even is a vector of progress in evolution! However, evolutionists never question that there is a naturalistic evolutionary process of some kind that explains the origin of all living things.
“Transitional” Fossils — The Missing Links
Evolutionists begin with the unquestioned assumption that evolution has occurred, starting with some primordial life form and progressing over time in a purely naturalistic way to produce all the kinds of living organisms on earth, past or present. Thus for “evidence” of evolution they need only to examine available fossils and attempt to arrange them in a sequence that appears to show progress over time. But a plausible sequential progression of intermediate stages is rarely, if ever, observed in the fossil record, which explains why we hear so much about “missing links.” Even Darwin himself was aware of this problem and said in his Origin of Species:
The number of intermediate varieties, which have formerly existed on the earth, [must] be truly enormous. Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely graduated organic chain; and this, perhaps, is the most obvious and gravest objection which can be urged against my theory.3
Why, indeed! For example, no one has observed progressive stages of “prebats” in the fossil record showing a mouse-like mammal gradually evolving into a bat with its long fingered wings. Evolutionists concede that what they consider to be the oldest bat fossils are 100 percent bats with some even showing evidence of sonar navigation.4 G.K. Chesterton put it simply: “All we know of the Missing Link is that he is missing — and he won’t be missed either.”
Many evolutionists now concede the dearth of transitional forms in the fossil record and feel obliged to come up with some sort of explanation for it. The late evolutionist Steven J. Gould bluntly admitted, “The extreme rarity of transitional forms in the fossil record persists as the trade secret of paleontology.”5
Again, Eldridge and Gould noted, “Most species during their geological history, either do not change in any appreciable way, or else they fluctuate mildly in morphology, with no apparent direction.”6
Gould even goes so far as to concede that not only are transitional stages not found in the fossil record, but in many cases we are not even able to imagine such intermediates:
The absence of fossil evidence for intermediate stages between major transitions in organic design, indeed our inability, even in our imagination, to construct functional intermediates in many cases, has been a persistent and nagging problem for gradualistic accounts of evolution.7
This conspicuous lack of fossil evidence for intermediate or transitional stages of evolution led Gould to a highly speculative rescuing hypothesis for evolution called “punctuated equilibrium,” or as it is sometimes called, the “hopeful monster theory.” In this scenario, the lack of fossil transitional forms is explained away by claiming that the transitional stages (hopeful monsters) being both unlikely and unstable occurred rarely and relatively quickly (on a geological time scale), leaving no fossil evidence. So what we actually see is stasis, i.e., no change over long periods of geological time!8 No wonder some evolutionists have argued that ancestor descendent relationships simply cannot be determined from fossils. For example, with regard to human evolution, Richard Lewontin said, “Despite the excited and optimistic claims that have been made by some paleontologists, no fossil hominid species can be established as our direct ancestor.”9
“Transitional” Living Organisms and Organs — Looking for the Dead Among the Living
When the fossil evidence fails to provide expected transitional stages, evolutionists often turn to living organisms in an attempt to arrange them in a way that appears to show a sequential process of evolution. An advantage of living organisms is that they allow the evolutionist to create an evolutionary scenario for the soft organs of the body. While we are becoming increasingly aware of evidence of soft tissue in fossils, most fossils show only hard tissue such as shells, teeth, and bones. Hard tissues represent a relatively small part of a living organism compared to their soft tissues. So with a bit of imagination, living organisms can sometimes be selectively arranged in a way to give the impression of an evolutionary sequence for soft tissue organs such as eyes, hearts, and kidneys.
In an effort to show evolutionary progress among living organisms, evolutionists look for structures or functions that appear to be intermediate in some way to those of other living organisms. These intermediate structures are then extrapolated to represent “transitional” stages in a sequential evolutionary progress. But while an organ or organism may be considered intermediate in appearance between two other organs or organisms, it does not necessarily mean that it represents an evolutionary transition between the two. Declaring something to be intermediate with regard to some arbitrary structure or character is merely an organizational decision, whereas declaring it to be transitional presumes an evolutionary or transformational process.
Living organisms are often used in an effort to explain the evolution of the eye. Darwin conceded in The Origin of Species that to suppose the eye could have evolved by natural selection “seems absurd in the highest degree,” and that to support his theory it would be necessary to demonstrate the existence of “numerous gradations” from the most primitive eyes to the most advanced ones. Since the fossil record provides no evidence for these gradations, evolutionists attempt to arrange the eyes of present-day living invertebrates and vertebrates into what appears to be a progressive evolutionary sequence. For example, in a journal devoted to giving evolutionary support for teachers, Lamb claims to have evidence from living hagfish that the vertebrate eye evolved through numerous subtle changes:
The great majority of the gradual transitions that did occur have not been preserved to the present time, either in the fossil record or in extant species; yet clear evidence of their occurrence remains. We discuss the remarkable “eye” of the hagfish, which has features intermediate between a simple light detector and an image-forming camera-like eye and which may represent a step in the evolution of our eye that can now be studied by modern methods.10
But a recent study of microRNA expression patterns in the hagfish and lamprey showed that the cyclostomes are closely related.11 This relation leaves evolutionists arguing whether the relatively simple hagfish eye is really a precursor of the more complex lamprey type eye or a degenerate form of that type of eye. From what then did the vertebrate eye evolve? There is a bewildering array of eyes found among the invertebrates. One of the world’s most distinguished experts on the eye, Sir Duke-Elder, said in volume one (The Eye in Evolution) of his monumental 15-volume work, System of Ophthalmology, that the eyes of invertebrates do not show a series of transitional stages:
The curious thing, however, is that in their distribution the eyes of invertebrates form no series of continuity and succession. Without obvious phylogenetic sequences, their occurrence seems haphazard; analogous photoreceptors appear in unrelated species, an elaborate organ in a primitive species (such as the complex eye of the jelly-fish Charybdea) or an elementary structure high in the evolutionary scale (such as the simple eyes of insects), and the same animal may be provided with two different mechanisms with different spectral sensitivities subserving different types of behavior.12
Duke-Elder was not even convinced that we ever will find a solution for the evolution of the eye:
Indeed, appearing as it does fully formed in the most primitive species extant today, and in the absence of transition forms with which it can be associated unless by speculative hypothesis with little factual foundation, there seems little likelihood of finding a satisfying and pragmatic solution to the puzzle presented by its (the eye’s) evolutionary development.13
With about 1.5 million named and categorized living species (and possibly several times more species unnamed or categorized), we might reasonably expect to see at least some evidence of a series of transitional stages among living organisms, but such is not the case. In his book Patterns and Processes of Vertebrate Evolution, evolutionist Robert Carroll concedes that very few examples of intermediate organisms or organs have been proposed:
Although an almost incomprehensible number of species inhabit Earth today, they do not form a continuous spectrum of barely distinguishable intermediates. Instead, nearly all species can be recognized as belonging to a relatively limited number of clearly distinct major groups, with very few illustrating intermediate structures or ways of life.14
“Transitional” Drawings and Illustrations — Making Your Own Data
When all else fails, there are always artists who will make a picture or model of any missing link the evolutionist might desire. Sadly, laymen are often strongly influenced by such fanciful illustrations. Consider the famous “March of Progress” monkey-to-man drawing commissioned by Time Life Books,15 one of the most famous and recognizable science illustrations ever produced. This drawing presumed to compress 25 million years of imagined human evolution into a row of progressively taller and more erect primates until finally a human walks away with a marine drill sergeant posture and gait.
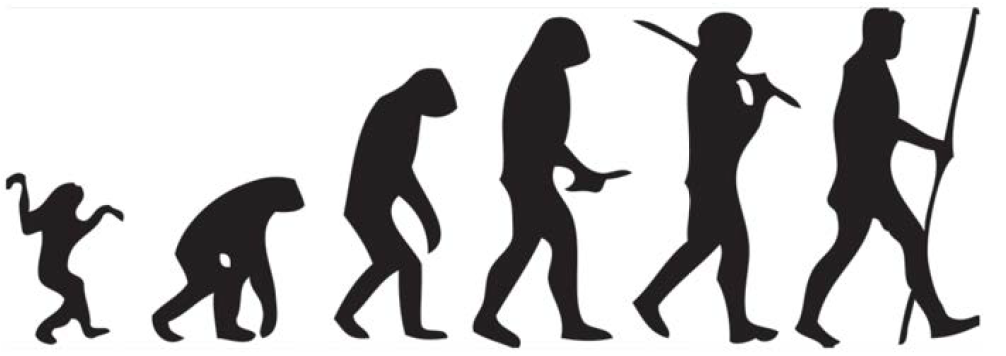
Many evolutionists have expressed their disapproval over this illustration showing a triumphalist linear progression of evolution that simply does not exist. Nonetheless, this “March of Progress” illustration has probably done more to convince uncritical laymen of the bestial origin of man than any other evidence.
Several years ago, the popular evolutionist Carl Sagan was on a television program where he showed a video clip of a rapid series of cartoon illustrations purporting to show amoeba-to-man evolution while a harpsichord solemnly played in the background. At the conclusion, the audience applauded enthusiastically, seemingly convinced that they had actually seen the whole sweep of amoeba-to-man evolution in a few minutes. We are living in an age where many are careless in distinguishing artistic license from scientific evidence.
But not all pictorial evidence for the imagined transitional stages of evolution is found in the popular literature meant for laymen. Imaginative drawings and illustrations are frequently found in the scientific literature intended for the specialist. An example of artistic license passing for “evidence” of transitional stages of evolution may be seen in efforts to explain the evolution of feathers.
Now that evolutionists are dead certain that dinosaurs evolved into birds (with many insisting that birds are in fact dinosaurs), they are left with the unenviable task of showing how reptile scales evolved into feathers. For years, evolutionists have insisted that feathers and scales are very similar, but nothing could be further from the truth.16 Scales are essentially continuous folds in the epidermis while feathers grow from individual follicles. These differences explain why the reptile must shed its entire skin to replace its scales while a bird sheds its feathers individually from feather follicles (in matched left-right pairs in the case of primary feathers). It is hard to imagine two cutaneous appendages more profoundly different than scales and feathers; they share almost nothing in common. In fact, feathers and their follicles show far more similarity to hairs and hair follicles than they do to reptilian scales, but there is no evolutionary scenario that relates the phylogeny of birds to mammals, so this fact is ignored by evolutionists. So evolutionists are stuck with making feathers out of scales, and to do so they must employ artists to illustrate transitional stages not seen in fossils or living creatures.
An attempt was made by Xu et al. to show the hypothetical stages of evolution from scale to feather.17 Their artist illustrates an elongated hollow scale first becoming a frayed or branching structure. This then somehow becomes a compound branching structure (see step II to IIIA below). To accomplish this, a structure with a simple branching pattern (all branching from one node) must implausibly become a compound branching structure (branching from several different nodes). The compound branching structure then undergoes another order of branching to give a superficial resemblance to a feather. Unmentioned is that in real life, all feather development must occur inside a follicle, where the feather is folded up in a sheath like a ship in a bottle. But then this presents no restrictions for an artist’s imagination and drawing.
What Do “Transitional” Stages Mean If There Is No Progress?
Can we have transitions or intermediates without progress? Many evolutionists are coming to the conclusion that there is neither purpose nor progress in evolution. In a recent survey of over 150 of the nation’s most influential and prestigious evolutionists (all members of the National Academy of Science), it was revealed that nearly 42 percent believe that evolution shows neither purpose nor progress.18 But if there is no purpose or progress in evolution, how can one identify incremental transitional changes in the process? Another 48 percent of these distinguished evolutionists believe evolution shows progress but no purpose. But how can there be progress without purpose? The English Wordnet dictionary defines progress as “an anticipated outcome that is intended or that guides your planned actions” and the Merriam-Webster dictionary defines progress as “a forward or onward movement as to an objective or to a goal.” Since nearly 80 percent of the evolutionists in the survey describe themselves as atheists, it is not surprising that they shun the notion of purpose in evolution. Purpose suggests the Creator (and accountability to the Creator), and that is unthinkable to these professional atheist/evolutionists.
Isn’t It Great to Be a Christian and Recognize God’s Purpose in Creation?
As Bible-believing Christians, we can gladly recognize the obvious that there is overwhelming evidence of intelligent design and purpose in God’s creation. Some evolutionists concede that they are aware of this evidence for design, but as the Bible says, they “suppress the truth in unrighteousness” (Romans 1:18). No better example of this suppression of the truth can be seen than the ardent atheist/evolutionist Richard Dawkins who wrote in the first page of his book titled The Blind Watchmaker: Why the Evidence of Evolution Reveals a Universe Without Design:
Biology is the study of complicated things that give the appearance of having been designed for a purpose.19
Dawkins concedes that this obvious appearance of design in biological systems cries out for some kind of explanation:
The complexity of living organisms is matched by the elegant efficiency of their apparent design. If anyone doesn’t agree that this amount of complex design cries out for an explanation, I give up.20
But the only explanation the atheist evolutionist can offer is that somehow nature “counterfeits” intelligent design. How sad.
The New Answers Book 4

Building on the previous New Answers Books, learn more about the Gospel and a young earth, death of plants and leaves, dragons, religious wars, cavemen, science, living fossils, and more.
Read Online Buy Book
Master Books has graciously granted AiG permission to publish selected chapters of this book online. To purchase a copy please visit our online store.
Footnotes
- Roger Lewin, “Evolutionary Theory Under Fire,” Science 210, no. 4472 (1980): p. 883– 887.
- D.L. Stern, “Perspective: Evolutionary Developmental Biology and the Problem of Variation,” Evolution 54, no. 4 (2000): p. 1079–1091; R.L. Carroll, “Towards a New Evolutionary Synthesis,” Trends in Ecology & Evolution 15, no. 1 (2000): p. 27–32; A.M. Simons, “The Continuity of Microevolution and Macroevolution,” Journal of Evolutionary Biology 15, no. 5 (2002): 688–701.
- Charles Darwin, The Origin of Species (1859; repr., New York: Avenel Books, Crown Publishers, n.d.).
- G. Jepsen, “Bat Origins and Evolution,” in Biology of Bats, W. Wimsatt, ed. (New York: Academic Press, 1970), p. 1–64; G.L. Jepsen, “Early Eocene Bat from wyoming,” Science 154, no. 3754 (1966): p. 1333–1339.
- Stephen J. Gould, “Evolution’s Erratic Pace,” Natural History 86, no. 5 (1977): p. 12–16.
- N. Eldredge and Stephen J. Gould, “Punctuated Equilibria: The Tempo and Mode of Evolution Reconsidered,” Paleobiology 3, no. 2 (1977): p. 145–146.
- Gould, “Is a New and General Theory of Evolution Emerging?” Paleobiology 6, no 1 (1980): p. 127.
- Stephen J. Gould, “The Return of the Hopeful Monsters,” Natural History 86 (1977): p. 22–30.
- Richard Lewontin, Human Diversity (San Francisco, CA: W.H. Freeman & Company, 1995), p. 179.
- T. Lamb, E. Pugh, and S. Collin, “The Origin of the Vertebrate Eye,” Evolution: Education and Outreach 1, no. 4 (2008): p. 415–426.
- Alysha M. Heimberg et al., “MicroRNAs Reveal the Interrelationships of Hagfish, Lampreys, and Gnathostomes and the Nature of the Ancestral Vertebrate,” PNAS 107, no. 45 (2010): p. 19379–19383.
- S.S. Duke-Elder, The Eye in Evolution, vol. 1, System of Ophthalmology (London: Henry Kimpton, 1958), p. 178.
- Ibid., p. 247.
- Robert L. Carroll, Patterns and Processes of Vertebrate Evolution (New York: Cambridge University Press, 1997), p. 9.
- F. Clark Howell, Early Man (New York, NY: Time-Life Books, 1965).
- David N. Menton, Formed to Fly, DVD (Hebron, KY: Answers in Genesis, 2007).
- X. Xu, Z. Zhou, and R.O. Prum, “Branched Integumental Structures in Sinornithosaurus and the Origin of Feathers” Nature 410, no. 6825 (2001): p. 200–204.
- G. Graffin, “Evolution and Religion,” The Cornell Evolution Project, http://www.polypterus.com/.
- Richard Dawkins, The Blind Watchmaker: Why the Evidence of Evolution Reveals a Universe Without Design (New York: Norton & Company, 1986), p. ix.
- Ibid.

Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- © 2024 Answers in Genesis