

Can Statistics Tell Us Whether Dinosaurs Had Feathers?
Recently, Dr. Gabriela Haynes published an article explaining why feathered dinosaurs are not viable and why creationists are unwise to accept the concept. Morphology-based statistical baraminology is one of the reasons some creationists do. Statistical baraminological analysis has been used to argue that dinosaurs had feathers.1 However, morphological statistical baraminology comes with significant baggage and numerous unwarranted assumptions.2
It is important at the outset to draw a distinction between baraminology proper and statistical baraminology. Baraminology, the study of created kinds, is a valid creationist area of research. Statistical baraminology is a subset of baraminology proper that uses statistical methods to try to determine the boundaries of created kinds. The majority of these studies are morphology based, although DNA-based studies are becoming more common. Other methods of baraminology exist, including hybrid studies—which is the biblical way to define kinds (Genesis 6:19, 8:17)—and the cognitum method,3 which Answers in Genesis used to complement hybrid studies when determining the ark kinds.4 Statistical baraminology, however, has been the predominant method in the field for the last two decades.
The first suggestion to use statistics to define kinds came in 1990.5 Prior to that, baraminology had been content with hybrid studies as proposed by the founder of baraminology, Frank Marsh.6 The call to use statistics was actualized in 1998 with a study on catarrhine primates.7 This study implemented something known as a baraminic distance correlation (BDC). This correlation is visualized as dij=mij/nij where d equals baraminic distance, m is the number of mismatched characters, and n is the total number of characters compared. While the distance is interesting, there is a fundamental assumption here—that similarity equals ancestry.
Creationists are undoubtedly familiar with the homology argument frequently advanced by evolutionists, namely that if two creatures look the same, they likely descend from a common ancestor. Fundamentally, the BDC equation makes the same assumption—that the more similar two organisms are, the more closely related they are. Obviously, this is false. Similar design might indicate common ancestry, but more commonly, it simply indicates a common design source. Yet the BDC equation makes an assumption that is antithetical to nearly a century of creationist arguments and remains the fundamental driving assumption of morphological statistical baraminology.
Reliable Data Source?
Like all other statistical measurements, statistical baraminology is dependent on its data. Yet almost all its data is derived from evolutionary phylogenetic data. That does not necessarily mean that it is bad or wrong, merely that it should be treated with skepticism. However, in statistical baraminology, the data is often treated as sacrosanct. Any question of bias is rejected unless it can be demonstrated.8 Yet the evolutionists themselves admit they go into their phylogenetic analysis with biases!9 Everyone has a worldview, and that worldview influences both their data collection and their data interpretation.
Even if the data is selected in an unbiased fashion, there is a further problem. The fundamental assumption of continuity and discontinuity is problematic. According to the statistical baraminology paradigm, organisms within a kind should share continuity with other members of the kind and be discontinuous with everything else. This discontinuity needs to be statistically significant and holistic. Without getting into the statistical weeds, statistical baraminology requires statistically significant results, which is highly dependent on character selection.10 Yet the characters are selected by evolutionists for the purpose of constructing a universal tree. Characters producing continuity will likely be favored by default. Since significance is so difficult to obtain, holism was emphasized for a while, but it too has been abandoned. Statistical baraminological literature effectively abandoned it by 2010 when a particular baraminology study attempting to broaden the human baramin moved Australopithecus sediba into the human baramin based purely on skull features.11 A similar agenda-driven paper moved Homo naledi12 into the human baramin based on the same features, and, though a later paper using more holistic data demonstrated it did not cluster with humans,1314 statistical baraminologists still consider H. naledi human.15
Importantly, morphology-based statistical baraminology has only been applied to humans once, and that was the first paper published in 1998.16 In that paper, the method could not separate apes and humans until over half of the data was thrown out. Since then, there has been no repeat. Yet it is not like data is unavailable to make the comparison. For example, two years before morphological statistical baraminology debuted, a paper was published comparing humans with modern apes and monkeys using 264 characters—17 apes and monkeys, humans, and 4 outgroups for a total of 22 taxa.17 Other papers are doubtless available as well.
If we grant all the premises of statistical baraminology and use the most up-to-date software, what results do we get? In no circumstance can we separate humans from the great apes. In both the baraminic distance graphs (Figures 1, 3) and the MDS plots (Figures 2, 4), humans cluster with the great apes.

Figure 1. Baraminic Distance Correlation graph of monkeys, apes and humans. The black squares represent significant continuity, the open circles represent significant discontinuity.
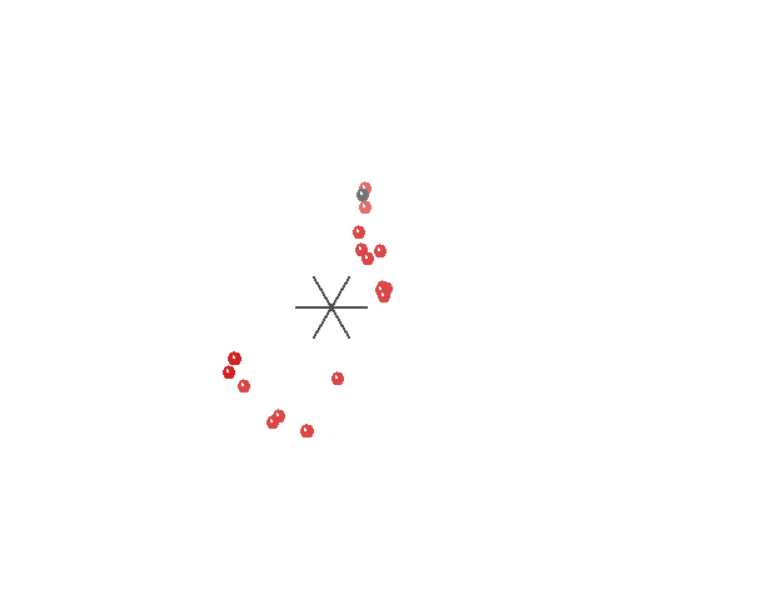
Figure 2. MDS plot of monkeys, apes and humans. The black dot is genus Homo (humans).
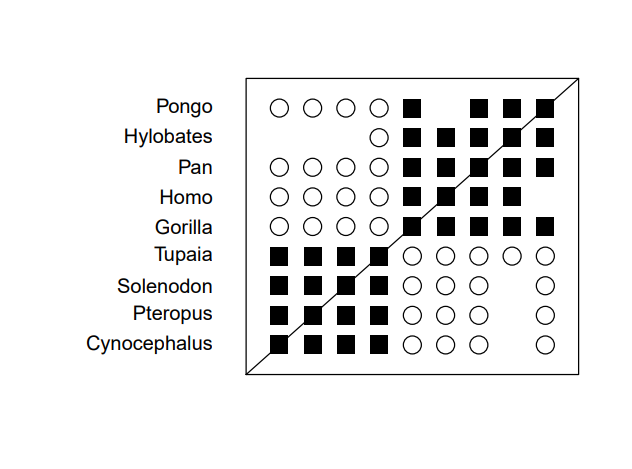
Figure 3. Baraminic Distance Correlation graph of apes and humans, with outgroups included.
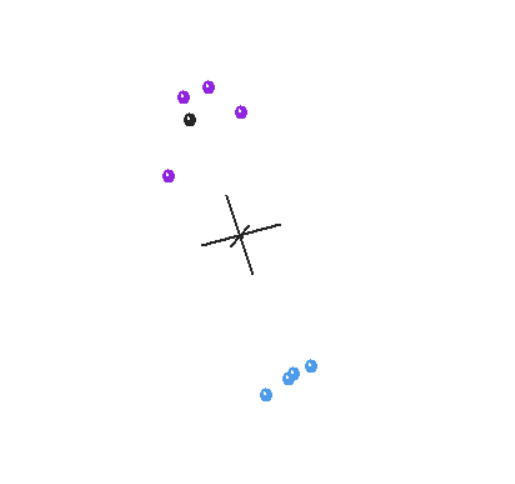
Figure 4. MDS plot of apes and humans. Black is genus Homo (human). The nearest dots represent the genera Gorilla and Pan (Gorillas and Chimpanzees).
This is a significant problem. If the data is reliable and the method works, as the advocates of statistical baraminology claim, we should be able to reliably separate apes and humans. After all, this is the one kind boundary we are clearly told in Scripture. Humans and apes share no ancestry. Yet, according to statistical baraminology, they are the same kind. Is the method at fault or the data—or both? Without more tests, we cannot be sure. It is likely the answer is partially both.
Creationists should carefully analyze any results obtained from statistical baraminology and ensure their results do not conflict with Scripture, known facts, and basic common sense before accepting them.
The larger issue here is not just statistical baraminology—though that is a major issue. The bigger issue is the importation of evolutionary assumptions into creation science, as well as unquestioning acceptance of what the evolutionists produce and attempts to incorporate it into the creation model. Like Dr. Haynes pointed out in her article, just because a creationist says it does not mean it is true or that we should agree with it. It is important to critically analyze everything, even if it comes from someone you think is on your side. Sadly, the lack of critical analysis is driving statistical baraminology and fundamentally driving a sect of creation scientists who have been labeled “young earth evolutionists.”18 One even argues that we should use the term evolution to talk about creation.19 Creationists should carefully analyze any results obtained from statistical baraminology and ensure their results do not conflict with Scripture, known facts, and basic common sense before accepting them.
Footnotes
- Matthew McLain, Matt Petrone, and Matthew Speights, “Feathered dinosaurs reconsidered: new insights from baraminology and ethnotaxonomy,” Proceedings of the International Conference on Creationism 8 (2018): 472–515, https://digitalcommons.cedarville.edu/cgi/viewcontent.cgi?article=1032&context=icc_proceedings.
- The content of this article is largely drawn from my article recently published in CRSQ: Harry Sanders and Matthew Cserhati, “Statistics, baraminology, and interpretations: a critical evaluation of current morphology-based baraminology methods,” CRSQ 58 (2022): 175–192, https://www.creationresearch.org/statistics-baraminology-and-interpretations-a-critical-evaluation-of-current-morphology-based-baraminology-methods.
- Roger Sanders and Kurt P. Wise, “The cognitum: a perception-dependent concept needed in baraminology,” The Proceedings of the International Conference on Creationism 5 (2003): 445–456, https://digitalcommons.cedarville.edu/cgi/viewcontent.cgi?article=1194&context=icc_proceedings.
- Jean Lightner, Tom Hennigan, Georgia Purdom, and Bodie Hodge, “Determining the Ark kinds,” Answers Research Journal 4 (2011): 195–201, https://answersresearchjournal.org/determining-the-ark-kinds/.
- Kurt P. Wise, “Baraminology: a young-earth creation biosynthetic method,” The Proceedings of the International Conference on Creationism 2 (1990): 345–360, https://digitalcommons.cedarville.edu/cgi/viewcontent.cgi?article=1456&context=icc_proceedings.
- Frank L. Marsh, “The Genesis kinds in the modern world,” Creation Research Society Quarterly 1, 30–38.
- D. Ashley Robinson and David A. Cavanaugh, “A quantitative approach to baraminology with examples from the catarrhine primates,” CRSQ 34, no. 4 (1998): 196–208.
- Todd Charles Wood, “Baraminology, the image of God and Australopithecus sediba,” Journal of Creation Theology and Science Series B: Life Sciences 1 (2011): 6–14.
- Mary P. Winsor, “The lessons of history,” Models in phylogeny reconstruction, eds. R. W. Scotland, D. J. Siebert, and D. M. Williams, (Oxford: Clarendon Press, 1994).
- Todd Charles Wood, Kurt P. Wise, Roger Sanders, and Neil Doran, “A refined baramin concept,” Occasional papers of the BSG (Baraminological Study Group) 3 (2003): 1–14.
- Todd Charles Wood, “Baraminological analysis places Homo habilis, Homo rudolphensis, and Australopithecus sediba in the human holobaramin,” Answers Research Journal 3 (2010): 71–90.
- Todd Charles Wood, “An evaluation of Homo naledi and “early” Homo from a young age creationist perspective,” Journal of Creation Theology and Science, Series B: Life Sciences 6 (2016): 14–30.
- Jean O’Micks, “Homo naledi probably not part of the human holobaramin based on baraminic re-analysis including postcranial evidence,” Answers Research Journal 9 (2017): 263–272.
- Jean O’Micks, “Further evidence that Homo naledi is not a member of the human holobaramin based on measurements of vertebrae and ribs,” Answers Research Journal 10 (2017): 103–113.
- Marcus Ross, “Evaluating hypotheses of hominin baraminology,” Proceedings of the Creation Research Society, 2022.
- Robinson and Cavanaugh, “A Quantitative Approach.”
- Jehekel Shoshani, Colin P. Groves, Elwyn L. Simons, and Gregg F. Gunnell, “Primate phylogeny: Morphological vs molecular results,” Molecular Phylogenetics and Evolution 5, no. 1 (1996): 102–154.
- Jake Hebert, “Is creation evidence ambiguous?” Institute for Creation Research, July 31, 2020, https://www.icr.org/article/is-creation-evidence-ambiguous.
- Ken Coulson, “Can a Christian believe in “evolution”?” New Creation Blog, August 7, 2020, https://newcreation.blog/can-a-christian-believe-in-evolution/.



Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- © 2024 Answers in Genesis