

Review: Your Inner Monkey
Your Inner Fish Episode 3
Your Inner Monkey, the final installment of Tiktaalik-discoverer Neil Shubin’s PBS miniseries Your Inner Fish, climbs to the top of the evolutionary tree of life to acquaint us with “our inner monkey” and show how an evolutionary past produced “the intelligent creatures with all our quirky flaws that dominate the planet today.”
Shubin’s case rests entirely on the assumption that all life evolved from a primitive common ancestor. As we examine his parade of proofs we shall see that it is this starting assumption, not the evidence in the human body as he contends, that determines his conclusions.
Heads I Win; Tails You Lose
Shubin travels back through presumed evolutionary history from skaters bruising their vestigial tails to monkeys swinging through trees. What about the tail? Does the human coccyx at the end of the vertebral column hang us in our rightful place on the tree of life? Or is the coccyx a perfectly designed structure that serves our needs as well as a monkey’s tail serves its need to swing in the trees? Do human embryos have a tail harking back to an evolutionary past
From the third to the seventh week of development the lower part of the human embryo’s curled, C-shaped body resembles a tail. Within this region many important things are happening, and none of those things have anything to do with swinging from trees, past or future!
Human embryonic development—like that of all vertebrate embryos—is organized by segments. In each segment are somites—blocks of cells that differentiate into muscles, bones, cartilage, and other supporting structures for that segment of the body or act as the scaffolding or stimulus for other structures to grow. In many cases, once an embryonic structure’s job is done, it regresses and disappears. (We discussed Meckel’s cartilage as an example of this in the review of Your Inner Reptile and another example—the urachus—in “Recapitulation Repackaged and Re-Applied.”) So it is with the so-called human embryonic tail.
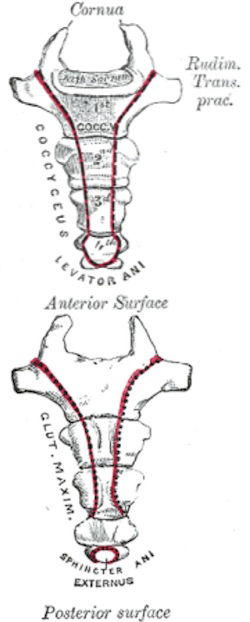
The coccyx, a set of small vertebrae at the bottom of the vertebral column, provides a secure anchor for the gluteus maximus and the muscles and ligaments that form the floor of the pelvis. Image: Gray’s Anatomy, Wikipedia.
Within this temporary so-called “tail” are a number of such somites, a secondary neural tube, and the lower part of the notochord. Cells in the notochord and secondary neural tube secrete signals that guide somite differentiation. But that’s not all! The secondary neural tube that forms in this “tail” is specifically responsible for forming the fibrous support for all those spinal nerves that extend out from the end of the spinal cord to innervate the lowermost parts of the body. When development is complete, the parts no longer needed disintegrate and leave the fibrous support for the nerves in place.1 The development of the support structures for these nerves is certainly an important event in the life of an embryo!
God designed humans to walk upright. He gave our coccyx several functions that make bipedality practical. First of all the gluteus maximus—the big muscle in the hip area—attaches to the side of the coccyx. Without the gluteus maximus we could not stand upright.2 Other muscles as well as several tough ligaments anchored securely to the coccyx form the pelvic floor, a series of slings supporting all the pelvic organs in their proper geometric orientation. Like anything else in this sin-cursed world, these supporting tissues can wear and weaken. When the pelvic organs begin to herniate downward, causing discomfort and even loss of control of certain bodily functions, we are reminded of the vital role the coccyx serves. The coccyx, essential to our vertical lifestyle, is not a marginalized evolutionary remnant on its way out but a vital part of our design.
A Handy Arrangement
Swinging from the trees to a fossil bed in Wyoming, Shubin introduces us to the lemur-like Notharctus. Like a lemur’s, the hand of Notharctus featured long fingers with fingernails (as opposed to the paws and claws of many other animals) and a thumb set at an angle to the other digits. This is the deepest human-like hand shape in the fossil record.
Such a hand is great for gripping the thinner branches of a tree. But what has this hand to do with human history? Nothing. Shubin maintains this primate made a handy evolutionary innovation that humans eventually inherited, but that interpretation is rooted in evolutionary presuppositions. Nothing in the study of living things shows such transitions from one kind of creature to another.
The human hand—perfectly suited for power and precision but without the extremely long fingers of an arboreal animal—shares some features with the lemur. Those common designs, however, are exactly what we expect from our Common Designer, the Creator God of the Bible, who made land animals and man on the same day, the 6th day of Creation Week about 6,000 years ago.
Walk Like a Man
Bipedality is considered the evolutionary essential for the human ancestral climb up the tree of life. We often hear that humanity’s ape-like ancestors came down from the trees after forests died and learned to walk upright. Shubin corrects this outdated idea which has been replaced by the notion that humanity’s ape-like ancestors were still at home in the trees while walking upright on the ground. The story changed when it became clear that the fossils evolutionists claimed for our ancestors were well-adapted for arboreal life.3
Evolutionists are not bothered by the fact that these fossilized apes cannot be made to walk with a human bipedal gait without dramatic contrivances.
While apes claimed as human ancestors seem well equipped for arboreal life, evidence for their supposed bipedality has a semantically checkered and even a somewhat contrived history. Australopithecus afarensis (Lucy)—the queen of supposed hominid bipedality4—has had her fossilized pelvis reconstructed in more than one way with drastically different results5—a fact not mentioned by Shubin or his Inner Monkey guest, Lucy’s discoverer Donald Johanson. The more these fossils are reconstructed and explored and their possible gaits analyzed—we say “possible” because the rocks have yet to produce a walking fossil for us to observe—the more we hear that they had a different sort of bipedal gait than humans. In other words, they had no genuinely bipedal gait at all.
Evolutionists are not bothered by the fact that these fossilized apes cannot be made to walk with a human bipedal gait without dramatic contrivances. They are committed to the idea that these apes were transitioning to human form anyway, so a presumed mosaic suits them. “Ardi” (Ardipithecus ramidus), from a bit deeper in the fossil record than Lucy, combines her “weird way of moving”6 with an opposable big toe and long fingers all set for climbing,7 making her Your Inner Monkey’s featured “mosaic.”
No mosaic gait will do for Your Inner Monkey’s Lucy, however. Computer graphics supply what nature did not, getting Lucy to take up her lumbering ape-body and walk with a perfectly modern human gait, strolling about Ethiopia’s Great Rift Valley. Artwork can’t change reality though. Living apes are able to achieve a bipedal gait, but it is not their habitual gait8 because they are not designed for efficient bipedality. Likewise, even if the australopithecine ape’s gait differed somewhat from that of chimps, on their best day they were not actually bipedal, hence the “concession” that these animals retained their arboreal adaptations. But even if Lucy and her cousins did have a more versatile anatomy than some other apes, tiptoeing through the jungle didn’t make anything ape-like turn into a person. Nothing could. God created Adam and Eve, the first humans, in His image the same day He created apes. We have His Word on it.
Back against the Tree
Is the human propensity for back problems a consequence of our evolutionary progress to bipedality, as claimed in Your Inner Monkey, or did Shubin and his guests put blame in the wrong place? Anatomist Bruce Latimer, whose opinions we discussed extensively in “Body Un-Designed?” calls the curves in the human back an “engineering nightmare.” For “proof” he re-enacted evolution, showing how difficult it is to “force” an animal’s vertebral bones into the “new position” they would occupy in an upright human and noting the intense pain and poor balance that would cause. Once again, the starting evolutionary assumption dictated the conclusion.
The curves of the human back, those we are born with and those that develop during infancy in response to our increasingly upright mobility, shift our center of balance over our feet and provide springy protection for the spine. The human back is designed for the way God designed us to walk. The fact that our bodies wear out does not link us to an evolutionary past. Back problems—like every other malady—are consequences of living in a sin-cursed world.

Opsins in the cone cells of our retinas absorb photons of light, change configuration, and signal the brain that a color has been detected. The different opsin molecules in humans (and also in Old World monkeys) absorb light from a different range of wavelengths. Because humans have all three opsins, we can detect a full spectrum of colors. Most animals have just two opsins and cannot distinguish between reds and greens very well. Image: from the video “Finding the Origins of Human Color Vision” at www.PBS.org
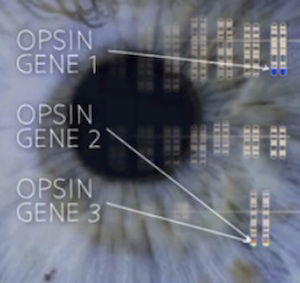
Two of our color opsin genes (#2 and #3) are located on the X chromosome. They detect medium and long wavelengths of light—the greens and the reds. Evolutionists believe that a primate ancestor evolved full spectrum color vision when opsin gene 2 (that detects medium wavelengths) duplicated itself and then mutated to become opsin gene 3, which absorbs longer wavelengths (redder colors). However, this interpretation is based on evolutionary belief that humans and all primates share a common ancestor. The presence of two similar genes (for medium and long wavelength opsins) side by side, as seen in humans and also in Old World monkeys, does not demonstrate evolutionary ancestry. Image: from the video “Finding the Origins of Human Color Vision” at www.PBS.org
Opsins for Color
Like those men and boys who must deal with the X-linked recessive condition commonly called “red-green color-blindness,” primates other than Old World monkeys have difficulty distinguishing red from green. In humans the problem is a gene rendered defective by mutation, but most animals simply lack one of the three genes9 that would be needed for full spectrum color vision.
Neither human color blindness nor the full spectrum color vision of most people demonstrates evolution from primate ancestors, but Shubin presents a different view. He says, “About 23 million years ago10 one group of primates evolved the ability to see many more colors.” He contends their visual spectrum widened when an opsin gene duplicated and then mutated. With full spectrum vision, the animal could tell which leaves and fruits were the most nutritious (so the story goes) and passed that beneficial trait on up the tree of life to humans.
The light-sensitive molecule God placed in the visual systems of most animals is called an opsin. We should not be surprised that our wise God chose to use this molecule, whose responsiveness to various wavelengths of light is readily altered by minor structural changes, in every animal that needs to detect light. (A form of this molecule is even found in some microbes.) Our retinal rods—which detect light but not color—contain an opsin. Cone cells in the retina are equipped with color-detecting opsins, and humans have three types, each responsive to different wavelengths. When an opsin molecule absorbs a photon of light, it changes shape, triggering a response interpreted by the brain as a color.
Two of the three color-sensitive opsins in humans are located on the X chromosome. Shubin indicates that because these genes are close together and similar in structure, they demonstrate our primate ancestry. But just as we see that our Creator used similar anatomical designs in diverse organisms, so we see He used many of the same sorts of molecules—including proteins and the genes that code for them—in diverse creatures. Common designs—genetic or anatomical—are exactly what we would expect to see in a world created as God described in His Word. (You can read more about opsins in “Perfect Molecule Shows Eyes Were Designed.”)
Brainy Background
As in Your Inner Fish and Your Inner Reptile, Your Inner Monkey uses both genetic and fossil-based arguments to support its claims. In a concluding segment to connect humanity back to an ancient worm, we meet amphioxus, a filter-feeding marine invertebrate that has a corresponding cousin in Cambrian rock.

This amphioxus (aka lancelet) specimen is Branchostoma lanceolatum. Amphioxus has a nerve cord with an enlarged area on one end but lacks the three-part brain seen in vertebrates. Amphioxus and vertebrates use similar genes to regulate nervous system development. Image: Hans Hillewaert, Wikipedia.
Amphioxus is an invertebrate whose ancestors are believed by evolutionists to have diverged from vertebrates over 520 million years ago. Amphioxus has a nerve cord with an enlarged area on the eating end, but it lacks the three-part11 brain architecture found in vertebrates. Similar genes regulate nervous system development in brainless amphioxus and govern division of a vertebrate embryo’s brain into three sections. Shubin claims “This means that the complex roots of our primate brains existed in the simpler creatures that first arose over 500 million years ago.”
A regulatory gene is like a switch. A switch can be used in many places. Its presence in this invertebrate and in vertebrates is not evidence of a shared evolutionary heritage but of a shared Creator.
Starting Points Determine Your Conclusions
Your Inner Fish has been a three-hour tour of the evolutionary story of our ascent up the tree of life. Shubin concludes, “There is something incredibly profound and I think beautiful in the idea that inside every organ, cell, and gene of our bodies lie deep connections to the rest of life on our planet.”
Having decided that all life must be explained without a Creator, evolutionists must believe increasingly complex creatures came into being through a series of interconnected events, even though such events are not seen in living creatures. Having already decided that all living things share a common evolutionary ancestry, they view embryologic development as a pattern providing evolutionary insight and interpret common designs as evidence for their position. The evolutionary starting point determines how they interpret comparative anatomy and genetics and essentially predetermines their conclusions.
The Bible, in the book of Genesis, records God’s eyewitness account of the six days in which He created all things. What we observe in anatomy, paleontology, and genetics is consistent with what we read in God’s Word. We understand the presence of common designs in a marvelously biodiverse world as the glorious expression of our Creator’s wisdom, power, and creativity. The human body, even with all the things that do go wrong with it in a sin-cursed world, is “fearfully and wonderfully made” (Psalm 139:14). When we get the starting points right, we can see the truth about our origins and better understand the rest of what the Bible tells us about our need for a Savior.
For more information:
- Review: Your Inner Fish
- Vestigial Hiccups, Folding Fish-eyes, and Other Fables: Our Fishy Forebears . . . Again!
- Organ Recital: A Parade of Presumed Design Flaws Devoid of Truth
- Not Ashamed of a Biblical Starting Point
- Scars of Evolution or of a Sin-Cursed World?
- Flexi-feet: Did Some Humans Fail to Leave Them in the Trees?
- Evolutionary Researchers: Human Hand Evolved for Dual Abilities
- Fossil Hand Turns Back the Clock on Tools for Human Evolution
- Did Ardi Head Up Human Evolution Before Lucy?
- Meet “Ardi”
- Evaluating the Gait Analysis of Ardi
- Life Lessons Illustrated in the Animal Kingdom
- Homo Erectus Had Skillfully Designed Tools
- ICR.org: Perfect Molecule Shows Eyes Were Designed
- Vestigial Organ Arguments: Useful or Unnecessary?



Footnotes
- Structures that serve vital but temporary roles in embryonic development may disappear in several ways. Whether by the mechanism of “apoptosis”—programmed cell death that seems to be active when a tight control on the final number of cells seems in order—or a simple disintegration as occurs with the leftover parts of the secondary neural tube, structures that have served their purpose in embryonic development are systematically eliminated, but this has nothing to do with the exhaustion of an earlier evolutionary role. D. Sapunar et al., “Morphological diversity of dying cells during regression of the human tail,” Annals of Anatomy 183 (2001): 217–222; and R.A. Nievelstein et al., “Embryonic development of the mammalian caudal neural tube,” Teratology 48, no. 1:21–31.
- It is the orientation of another gluteal muscle, the gluteus medius, that makes humans but not apes able to have a truly bipedal gait. See the photographs in “Lucy, the knuckle-walking ‘abomination’” to learn more.
- Read more about it in “Lucy’s Ups and Downs,” “Lucy in the Trees,” and “A Look at Lucy’s Legacy.”
- Read more about Lucy’s locomotion in “A Look at Lucy’s Legacy.” and “Lucy, the knuckle-walking ‘abomination’”
- After CT analysis of Lucy, for instance, evolutionist Christine Berge wrote, “The results clearly indicate that australopithecine bipedalism differs from that of humans. (1) The extended lower limb of australopithecines would have lacked stabilization during walking; and (2) the lower limb would have shown greater freedom for motion, which can be interpreted as the retention of a partly arboreal behavior” (from C. Berge, “How did the australopithecines walk? A biomechanical study of the hip and thigh of Australopithecus afarensis”. Journal of Human Evolution 26, no. 4:259–273, www.sciencedirect.com/science/article/pii/S0047248484710165, quoted in “A Look at Lucy’s Legacy.”) Australopithecus sediba is described as having a particularly awkward foot-rolling sort of gait, “sashaying” with a “swag” in the words of its paleontological parents. The team that discovered Ardi (Ardipithecus ramidus) described the best bipedal gait they could achieve in their analysis as a “weird way of moving.” (from National Geographic, quoted in “Did Ardi Head Up Human Evolution Before Lucy?”) As with the other candidates for human ancestor, Ardi’s gait analysis failed to provide Ardi with a truly bipedal gait.
- National Geographic, quoted in “Did Ardi Head Up Human Evolution Before Lucy?”
- Ardi’s canine teeth seem small for an ape, a trait described as another human-like mosaic quality on Your Inner Monkey but also attributable to simply being female. Apes typically have a great deal of gender-based dimorphism. In any case neither teeth nor toes make this ape anything but another variety of extinct ape.
- See the photographs in “Lucy, the knuckle-walking ‘abomination’.”
- Interestingly, while three different opsins—L, M, and S—are needed for trichromatic color vision, animals that are “missing” one of these opsins are not all missing the same ones. L, M, and S stand for the long-, medium-, and short-wavelength colors to which each opsin in maximally responsive.
- In his book Your Inner Fish, Shubin reports a different number, 55 million years ago, but what’s a few million years between our inner monkeys?
- Vertebrate brains are organized into three main divisions, loosely termed the forebrain, the midbrain, and the hindbrain.







Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- © 2025 Answers in Genesis