
The Beauty of the Peacock Tail and the Problems with the Theory of Sexual Selection
Originally published in Journal of Creation 15, no 2 (August 2001): 94-102.
Abstract
According to the theory of sexual selection, the peacock tail has gradually evolved because the peahen selects beautiful males for mating.
The peacock tail contains spectacular beauty because of the large feathers, bright, iridescent colours and intricate patterns. The colours in the tail feathers are produced by an optical effect called thin-film interference. The eye pattern has a high degree of brightness and precision because the colour-producing mechanisms contain an extremely high level of optimum design. According to the theory of sexual selection, the peacock tail has gradually evolved because the peahen selects beautiful males for mating. However, there is no satisfactory explanation of how the sexual selection cycle can start or why the peahen should prefer beautiful features. In addition, there is irreducible complexity in both the physical structure of the feather and in the beautiful patterns.
Most birds have two types of tail feather: flight feathers and tail-coverts. The flight feathers provide stability during flight, while the tail-coverts ‘cover’ and protect the tail region. In the vast majority of birds, the tail-coverts are small feathers, just a few centimetres long. However, some birds like the peacock have very large tail-coverts for decorative purposes. These decorative feathers are also referred to as ornamental feathers, or display feathers.1 It should be noted that a peacock is a male peafowl and a peahen is a female peafowl. The peahen does not have any decorative feathers.
When a peacock displays his tail feathers during courtship, a magnificent ‘fan formation’ of feathers forms a beautiful backdrop to the body of the peacock as shown in Figure 1 (below). An adult peacock has an average of 200 tail feathers and these are shed and re-grown annually. Of the 200 or so feathers, about 170 are ‘eye’ feathers and 30 are ‘T’ feathers. The ‘eyes’ are sometimes referred to as ocellations.
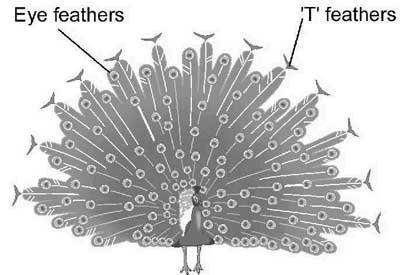
Figure 1. Peacock with tail feathers displayed.
This paper describes some of the complex structures that are responsible for producing the beautiful features and why the beauty of the peacock is evidence for intelligent design. The paper also describes the theory of sexual selection and shows that there are serious problems with the theory.
Fan formation of displayed feathers
When the peacock feathers are displayed there are several beautiful features that can be seen:
- Fan formation of feathers
- Uniform distribution of ‘eyes’
- Intricate ‘eye’ feathers
- Intricate ‘T’ feathers
One reason for the beauty of the displayed feathers is that they form a semi-circular fan over an angle of more than 180 degrees. The fan formation is very even because the axis of every feather can be projected back to an approximately common geometrical center. The radial alignment of feathers requires the root of each feather to be pointed with a remarkable degree of accuracy. Another remarkable feature of the displayed feathers is that they are ‘deployed’ into position by muscles in the peacock’s tail. Not only can the peacock deploy the feathers, but he can also make them vibrate and produce a characteristic hum.
Another beautiful feature of the displayed feathers is the uniform spacing of the eyes. Even though the display contains around 170 eye feathers, they are all visible and all spaced apart with a remarkable degree of uniformity. All the eyes are visible because the feathers are layered with the short feathers at the front and the longer feathers at the back. The eyes have an even spacing because each feather has the right length.
Each ‘eye’ feather and ‘T’ feather is an object of outstanding beauty in itself. The eyes contain beautiful patterns, and the ‘T’-shaped feathers form a beautiful border to the fan.
The eye feather
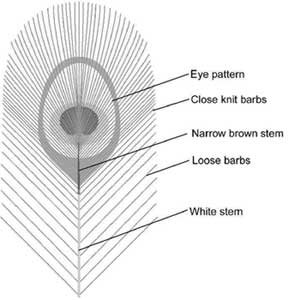
Figure 2. Structure of the eye feather.
Figure 2 (right) shows a sketch of the top section of the eye feather. There are several beautiful features to the feather:
- Bright colours
- Intricate eye pattern
- Loose barbs below the eye pattern
- Absence of stem in the top half of eye pattern
- Narrow stem in the bottom half of eye pattern
- Brown coating of the stem near the eye pattern
The bright colours and intricate shapes of the eye pattern are the most striking aesthetic features. The loose barbs on the lower part of the feather are beautiful because they make a contrast with the neatness and precision of the barbs in the eye pattern.
The last three features in the list above are usually only noticed by very careful observers. However they represent important ‘finishing touches’ which make an important contribution to the beauty of the feather. The absence of a stem in the top half of the eye is an important detail because it prevents the pattern from being divided into two sections. The stem is not needed because the barbs fan out around the top of the feather. The narrowness of the stem in the bottom half of the eye pattern is important because this makes the stem fairly obscure. The stem can be narrow because it has a deep section in the area of the eye pattern. The brown coating of the stem in the area of the eye pattern is very important because the stem is a natural white colour and this would be too conspicuous for the eye pattern. It is interesting to note that the stem is white everywhere except local to the eye pattern. This strongly indicates that the brown coating near the eye pattern is a deliberate feature.
A large eye feather has been examined at Bristol University to determine the number and size of each part of the feather. The number and size of barbules was estimated by examining sample sections of barbs with a microscope. The data for the feather are summarized as follows:
- Length of feather = 1.3 m
- Number of barbs = 290
- Maximum length of barbs = 200 mm
- Average length of barbs = 105 mm
- Barbules per mm on one barb = 32 (16 each side)
- Length of barbules in eye pattern = 0.8 to 1 mm
- Length of barbules below eye pattern = 2 to 3 mm
- Total number of barbules in feather = nearly 1 million
The results show that a large peacock tail feather is very large both in terms of size and number of barbules. The unique length and structure of the peacock display feathers is acknowledged by bird experts.2,3
The colours in the eye feather
The colours in the peacock tail are particularly beautiful because they are bright and iridescent. An iridescent colour is a colour that changes with the angle of view. The colours are not produced by pigments but by an optical effect called thin-film interference that takes place in the barbules.4 In technical terms, the peacock has ‘structural colours’.
In the eye pattern, the barbules appear bronze, blue, dark purple and green. Away from the eye region, the barbules are uniformly green. The colours in the eye feather can only be seen on the front surface of the feather because this is where the barbules are positioned. The back of the feather is uniformly brown because the barbs contain a brown pigment. To understand how thin-film interference is produced in the peacock tail, it is first necessary to understand the detailed structure of the feather.
Structure of the barbules
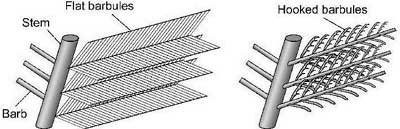
Figure 3. Peacock barbules.
(a) Tail feather barbules (b) Flight feather barbules
The basic structure of the peacock tail feather in the eye region is shown in Figure 3(a) (right). For comparison, the structure of a typical flight feather is shown in Figure 3(b) (right). Like the flight feather, the peacock tail feather has a central stem with an array of barbs on each side. Also, individual barbs have an array of barbules on each side of the barb. Even though there is a basic similarity with a flight feather, the peacock tail feather has an unusual barbule structure. The barbules are like long flat ribbons that overlap to form a flat surface on top of the barbs. (Under a microscope the barbules are actually slightly curved and segmented and the surface has a bubbly appearance). In contrast, a flight feather has narrow barbules which do not cover the barbs. Other types of birds such as hummingbirds, pigeons and kingfishers have some patches of flat iridescent barbules, but the peacock has the largest iridescent barbules of any known bird.5
The colours of the barbules dominate the front face of the tail feather because they completely cover the barbs. The barbules are not very visible from the back of the feather because the barbs are quite close together.
Thin-film interference in the barbules
Thin-film interference can be produced in one or more layers of a very thin and transparent material. Usually the thin film is placed on a dark surface. The thickness of the transparent material must be close to the wavelengths of visible light. Visible colours have wavelengths between 0.4 and 0.8 µ and thin films typically have a thickness of between 0.3 and 1.5 µ. Another requirement for thin-film interference is that the thin film must have a refractive index that differs from air so that the light is retarded when it passes through the thin film. Thin-film interference commonly occurs in oil slicks on a wet road. The oil will often form a thin layer on the wet surface of the road or on the surface of a puddle, the thin-film producing blue and green colours.
In the case of the peacock, thin film interference takes place in three layers of keratin which cover the barbules as shown in Figure 4. Each barbule is about 60 µ wide and 5 µ thick.6 The barbules have a foam core that is 2 µ thick and this is covered with three layers of keratin on each side, as shown in Figure 4 (below). The keratin layers are very thin, being about 0.4–0.5 µ thick.7
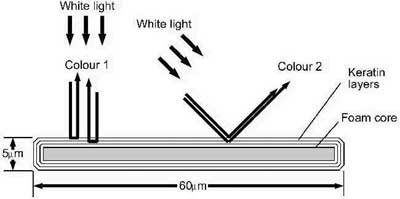
Figure 4. Cross-section of peacock barbule.
The principle of thin-film interference in a single layer of keratin is shown in Figure 4. White light is reflected off the front and back surfaces of the thin film. The light which passes through the keratin is retarded and therefore when it emerges from the keratin, some of the colour components of white light are out of phase with the light-waves that were reflected from the front surface. When two wave trains of the same colour are out of phase, destructive interference removes the colour. In the case of white light, the result of the interference is a reflected colour due to the remaining colour components of white light. In practice, interference occurs simultaneously in all three thin films.
The only pigment in the peacock tail is melanin, which gives the barbs a uniform brown colour. This provides a dark background colour for the thin-film interference in the keratin layers. The different colours in the eye pattern result from minute changes in the depth of thickness of the keratin layers.8 In order to produce a particular colour, the keratin thickness must be accurate to within about 0.05 µm (one twenty thousandth of one millimetre!).
The barbules in the peacock feather contain a high degree of optimum design. The thickness of the keratin layers is optimal for producing the brightest thin-film colours. The dark brown background colouring is optimal because it prevents light shining through the back of the feather. The three layers add to the brilliance of the colours in the feather by adding multiple components of light. The barbules are also slightly curved in the longitudinal direction.9 This curvature causes a mingling of slightly different colours, which produces a softening of the colours seen in the keratin layers.10
The eye pattern
The particular beauty of the eye pattern comes from the rounded shapes that have a high degree of resolution as shown in Figure 5 (below). The ‘pupil’ of the eye is formed by a dark purple cardioid and the ‘iris’ is formed by a blue ellipsoid. These shapes are located within a pointed bronze ellipsoid that is surrounded by one or two green fringes.
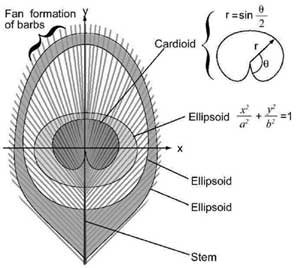
Figure 5. Mathematical curves in the eye pattern.
A very important feature of the eye pattern is that it is a digital pattern which is formed by the combined effect of many thousands of individual barbules. Some patterns in nature are formed by natural growth mechanisms, as with the spiral shape of the nautilus shell. However, the eye pattern in the peacock tail requires the precise coordination of independent barbs and this cannot be achieved by a simple growth mechanism. Barbules on adjacent barbs coordinate perfectly with each other to produce the eye pattern.
The spacing of colours on each barb must be specified by instructions in the genetic code. To specify the pattern, there must be timing or positional instructions in the DNA which causes the right thickness of keratin to be grown on the right barbule and on the right barb. To help appreciate the precise nature of the information in the genetic code, it is helpful to consider the mathematical complexity involved in calculating the required spacing of colours on each barb.
Required colour spacing on barbs
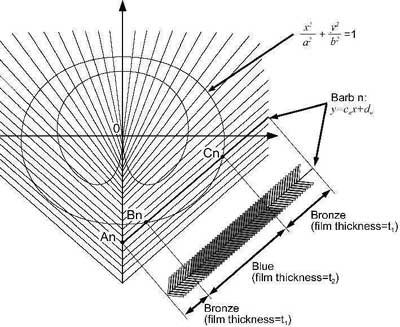
Figure 6. Intersection points on barbs.
Figure 6 (right) shows the colour spacing on a single barb. Along the first part of barb ‘n’, the thickness of the keratin films on the barbules gives a bronze colour. Then an abrupt and minute change in thickness of the keratin films produces a blue colour. Another abrupt and minute change in thickness of the keratin films so produces a bronze colour. The abrupt nature of the changes in thickness is important because if the changes were gradual then there would be a gradual change in colour.11 The abrupt changes in thickness of keratin along a barb are an amazing feature because it involves sudden and precise changes in the dimensions of the barbule. Even more amazingly, along the length of the barb the thickness of the keratin does not continually get thicker and thicker (or thinner and thinner) but it involves both increases and decreases in thickness.
The required length of the colour sections on each barb can be determined mathematically by finding the points where the barbs intersect with the curves as shown in Figure 6. For example, to find the positions of the points Bn and Cn the following procedure can be followed. Firstly the equation for the ellipsoid (conic function) can be written as:
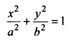 (1)
(1)
Then the equation for the straight line of barb n can be written as:
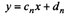 (2)
(2)
By taking the equation for the ellipsoid (1) and substituting it into the equation for a straight line (2) and eliminating y we get an equation for x as follows:
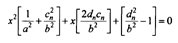 (3)
(3)
This equation can be solved as a quadratic to get two solutions for x, the two intersection points xB(n) and xC(n). The y coordinates of these points then can be found from either (1) or (2). Then, the length of the first section of bronze colour on barb n can be found by geometry:
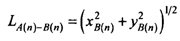 (4)
(4)
A similar procedure can be used for the intersection points on the cardioid shape and the outer green fringes. For each barb there are on average about four points at which colour changes and so there are on average four positions to calculate. Since there are around 50 barbs on each side of the pattern and since every one of these barbs has a unique spacing of colour, it is necessary to calculate 200 intersection points in order to construct the whole eye pattern.
‘T’ border feathers
The long ‘T’ border feathers provide a beautiful border to the tail feathers because they form an inverse shape to the peacock eye as shown in Figure 7 (below). An inverse shape is beautiful because the inside profile of the T feather follows the outline of the eye pattern. The T feathers often form an ‘ogee’ curve on each side of the feather as shown in Figure 7. An ogee curve is beautiful because it is both concave and convex. For this reason, ogee curves are used in architecture in structures such as arches. The formation of an ogee curve from individual barbs is yet another remarkable feature of the peacock tail. Each barb at the end of the T feather has a unique length and curvature and all the barbs coordinate exactly with each other to form the curved T.
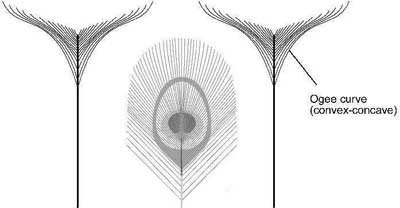
Figure 7. The 'T' feathers and 'eye' feathers.
Information content in the genetic code
Every detail in the peacock tail must be defined by genes in the genetic code of the peafowl. Since the tail feathers have very complicated structures and colour-producing mechanisms, there must be a large amount of design information in the genetic code.
It is difficult to determine how many genes would be required to specify the aesthetic features of a peacock tail feather because it is not known how the tail feather grows. However, a conservative estimate can be made by assuming that each separate aesthetic feature is specified by one gene. By assuming that each colour and each shape within the eye pattern represents a separate feature, and taking into account the other features discussed in this paper, the total number of aesthetic features in a single feather comes to about 20. Therefore an estimated 20 genes are required for the peacock tail. This may be a very conservative estimate. In particular, it may be that many genes are required to produce each shape in the eye pattern since the eye pattern is formed from the coordinated arrangement of over 100 barbs. In addition, the fanning-out of barbs in the top of the feather, where there is no stem, is a complex feature that may well need several controlling genes.
Even if only 20 genes are required to specify the beautiful features of the peacock tail, this still amounts to a lot of genetic information. A gene typically consists of 1,000 chemical units of information (base pairs). Therefore, 20 genes would contain many thousands of chemical units of information. According to evolutionists, all of this information has appeared gradually by genetic mistakes and by sexual selection.
The theory of sexual selection
The theory of sexual selection was first proposed by Charles Darwin in The Descent of Man.12 Even though this theory has always been controversial, most evolutionists now believe that it can explain how beautiful features could evolve from nothing.13
According to sexual selection, a female can have a preference for a mate with a feature such as a long tail. Over a long period of time, sexual selection is believed to be able to develop a particular feature to a great extent. For selection to work, a number of things are thought to be typically required. Firstly, the male must have an aesthetic feature. Secondly, the female must have a preference for that particular aesthetic feature. Thirdly, the female must be able to have the opportunity to view a number of different males before mating. Fourthly, the female must be able to have some control over which male mates with her.
Sexual selection is a circular process based on a particular fashion. When females have a preference for a long tail, the selection of a male with a long tail is an advantage because the male offspring will have long tails and therefore be more successful at mating. A key aspect of sexual selection is that ‘fitness’ is not measured in ability to escape from predators but in ability to produce offspring. Evolutionists fully recognize that sexual selection would often produce features that reduce the ability to escape from predators because aesthetic features often make a creature more conspicuous and slower. However, if females prefer beautiful males for mating, then the advantage of beauty can outweigh the advantages of camouflage and maneuverability. According to the theory of sexual selection, ornamental features will develop to the point at which the disadvantages of being caught by a predator outweigh the advantages of being selected by a female.14
Evolutionists recognize that a female such as a peahen does not have aesthetic appreciation and that the preference of the female is based on an instinctive response. In addition, it is recognized that the instinctive response needs to be specified by one or more genes in the genetic code15 called preference genes.
Do the peacock tail feathers play a role in the courtship ritual?
There is no doubt that the peacock tail feathers do play a role in the courtship ritual of peafowl. Many creatures have a courtship ritual that acts as a cue for mating. In the case of the peafowl, the peacock shows his intention to the peahen by displaying his feathers. However, even though the display feathers have a role in the courtship ritual, this does not necessarily mean that the female is ‘attracted’ to the feathers. When the peahen observes the feathers of the peacock, it may be that her only reaction is to understand that the peacock is ready for mating.
Of course, the beauty of the peacock tail display is vastly beyond what is required to make a cue for the peahen. However, it is the Creator’s prerogative to design a cue that is vastly more complicated than what is required. One could argue that the courtship ritual is an appropriate place to install great beauty because courtship is a beautiful process in itself.
Do preference genes exist in the peahen?
Biologists have carried out studies on the behavior of peafowl during courtship to try to determine if the peahen is really attracted to particular features of the peacock. One study has revealed that peahens do recognize obvious features in the peacock such as the number of eye feathers.16 The results of this study indicated that the peahen prefers males with a greater number of eyes. However, other studies have indicated that the peahen has little or no interest in the appearance of the peacock.17 There is no evidence that peahens can recognize subtle aesthetic features.
If there is a preference gene for aesthetic features, this does not prove that the sexual selection theory is true.
If there is a preference gene for aesthetic features, this does not prove that the sexual selection theory is true. The reason for this is that the Creator may have installed a preference gene as a means of ‘maintaining’ beautiful features. Beauty generally gives a disadvantage in terms of escaping from predators. If a peacock lost its colours due to a gene mutation, it would suddenly find itself more protected from predators. This is an example of where a loss of information could be a great advantage in terms of survival. Therefore, it is conceivable that the Creator would deliberately create preference genes for prominent aesthetic features such as colour.
Even in the case of subtle aesthetic features, it is conceivable that the Creator may have created preference genes in order to root out genetic mistakes. However, there would be less selective pressure for subtle features to be lost since they do not affect the ability of the peacock to escape from predators.
At present, there is no conclusive evidence about the existence of preference genes. Future experiments in this area should be very interesting, especially if a preference gene could be directly identified in the genetic code. It is possible, though, that the peahen does have preference genes for obv If future experiments show that there are no preference genes in the peahen, then the theory of sexual selection would absolutely collapse. However, even if future studies do reveal a preference gene for obvious aesthetic features, there are still some very serious problems with the theory of sexual selection. Five of the problems are: When females have a preference, that preference becomes self-perpetuating.18 However, there is no reason a fashion should always be a ‘beautiful’ fashion. According to evolution, preference genes appear by totally random processes and therefore there could be a fashion for all kinds of features including ugly features. In reality, where males have decorative features, such as the birds of paradise and the peafowl, it is clear that every aesthetic feature contains a very high degree of aesthetic merit. To overcome the problem that females always prefer beautiful features, evolutionists have proposed the ‘good genes’ theory that proposes that beauty is directly related to health and fitness.19 However, the decorative features found in nature are so overwhelmingly beautiful that it would require an extremely strong correlation between beauty and health and there is no evidence for such a strong correlation. Another big problem with the theory of sexual selection is the question of how the sexual selection cycle can start by chance. The cycle cannot start until there is both a trait gene and a preference gene. Therefore for a sexual selection cycle to get started there must be the appearance of two new genes in the DNA. Since genes contain complex information and since the preference gene and trait gene are useless on their own, it must be concluded that sexual selection could never spontaneously commence on the basis of incremental changes to the DNA. To overcome the problem of the simultaneous appearance of two new genes, evolutionists have proposed that the two genes appear at different places and different times in the following way.20 First, a female spontaneously produces a preference gene for a male with, say, a long tail. This gene lies dormant for perhaps many generations without any opportunity to be expressed. Then one day, a male spontaneously generates a gene which produces a longer tail. The female then selects that male and some of their offspring have both the trait gene and the preference gene. Therefore, the cycle is in place and ready to develop and perpetuate long tails. At first, this scenario may seem plausible. However, it still relies on simultaneous chance events. Firstly, there must be a preference gene that matches a trait gene. Secondly, there must be a chance meeting between the right female and male. The first gene to arise also has to survive genetic drift until the male gene arises. Therefore, even with the scenario given by the evolutionists, it is clear that the sexual selection cycle is extremely unlikely to get started. The starting of one sexual selection cycle is difficult to explain by chance. However, when a creature contains many separate aesthetic features, the problem becomes even more pronounced because many cycles must be started. In the case of the peacock, there are many aesthetic features in the tail. In addition, the peacock also has several aesthetic features in the rest of its body. For example, it has a bright blue neck, patterns around the eyes, a crown on the head and speckled contour feathers. This array of features would probably require many sets of preference genes and trait genes. It may well be possible that a peahen has a preference for obvious features such as a long tail. However, there are some extremely subtle features in the peacock which are not easy to recognize. These subtle features include an absence of a stem in the upper part of the eye pattern, the brown colouring of the stem near to the eye pattern and the intricate shape of the ‘T’ feathers. It may be reasonable to argue that a peahen could recognize whether a peacock had lost its eye feathers or T feathers. However, to discern subtle changes in these feathers would require tremendously detailed observation. The above features are so subtle that many people do not notice them. In addition, it is necessary to get quite close to the feather to recognize such features. Since peahens do not undertake close visual inspections of the peacock, they would have to have a much better eye for detail than a human being in order to recognize the subtle features of the peacock tail. Darwin himself recognized the problem of subtle aesthetic features. Darwin said, ‘Many will declare that it is utterly incredible that a female bird should be able to appreciate fine shading and exquisite patterns. It is undoubtedly a marvellous fact that she should possess this almost human degree of taste’.21 What is really incredible is that evolutionists really believe that a peahen is able to recognize fine shading and exquisite patterns. There is no evidence that the peahen can recognize such subtle aesthetic features. Some of the structures that produce the aesthetic features in the peacock tail are irreducible. This means that they require several features to be simultaneously present in order for the structure to function. One example of an irreducible structure is the thin-film interference. Thin-film interference in a feather requires all of the following features to be simultaneously present: Since evolution is supposed to work by changing one parameter at a time, thin-film interference cannot be produced by a process of evolution. For example, if there was a random gene mutation that suddenly caused a barbule to become flat, this change would not be enough to cause thin-film interference. Even if a barbule were to become flat and acquire a layer of keratin, this would still not produce a thin-film colour unless the keratin was the right thickness. Getting the right thickness of keratin by chance is very difficult because the keratin thickness has to be within a very narrow range for thin-film interference to work. For thin-film interference to work, the thickness of keratin normally has to be within a range of 0.4–1.5 µ. However, keratin can be formed in thicknesses from 0.2 µ up to 1 mm. For example, nails and feather stems have keratin with a thickness of around 1 mm. If one considers 1,000 different layers of keratin which all have a different thickness ranging from 1 µ, 2 µ, 3 µ, etc., all the way to 1,000 µ (1 mm), only one or two out of the thousand thicknesses would produce thin-film interference. Therefore, it is inconceivable that a peacock could acquire a flat barbule and exactly the right thickness of keratin simultaneously. The only way to produce iridescent feathers is to make a fully functioning flat thin-film barbule from the beginning. The fact that thin-film interference is a delicate and sophisticated mechanism is fully acknowledged by evolutionists. For example, Mason says the following: This is an important quote because Mason’s studies on the colour of peacock feathers are referred to by most modern texts on bird coloration. Notice how the author refers to the ‘perfection of nature’s method’, and marvels at how the thin-film is maintained in successive generations. If it is hard to understand how the peacock ‘maintains’ its delicate structures through successive generations then how does the evolutionist explain how it could have evolved in the first place? According to evolution, a complex pattern like the eye pattern in the peacock’s feather has evolved by the accumulation of hundreds of genetic mistakes occurring over vast periods of time. However, patterns like the blue ellipsoid in the eye are irreducible, i.e. they require several features to be simultaneously present in order for there to be a clear pattern. If only one barb in a peacock tail feather was to have a patch of blue colour this would not produce a beautiful pattern. Such a random change would arguably cause the peahen to deselect, not select the pattern. Since evolution requires every step change to have a selective advantage, the eye pattern cannot evolve but must be designed complete from the beginning. The difficulties with the theory of sexual selection have led some evolutionists to develop alternative theories for the origin of beauty. The existence of these alternative theories suggests that the theory of sexual selection is not sound. The main alternative theories are:Problems with the theory of sexual selection
(i) Why should the female select a ‘beautiful’ feature?
(ii) How can the sexual selection cycle start by chance?
(iii) How can multiple aesthetic features start by chance?
(iv) How can the female appreciate subtle features?
(v) Some features contain irreducible mechanisms
‘The theory of thin films as the cause of iridescence, although it fits all the observed facts, cannot but inspire one to marvel at the perfection of nature’s method of producing these colours with such uniformity through successive generations, especially when a slight general variation in thickness of the films of the feathers of a bird, such as a peacock, would be enough to alter its coloration completely.’22
(vi) Some features contain irreducible beauty
Alternative theories for the existence of beauty
(i) Male pecking order
Some evolutionists believe that males like the peacock compete with other males in order to win a privilege of mating with a female.23 It is believed that the competition can be based on the beauty of a display. The idea is that the male with the most impressive display frightens the other males into submission. A major problem with this theory is that it cannot explain why there should be subtle aesthetic features.
(Ii) Camouflage
Some evolutionists claim that the peacock tail gives a camouflage advantage.24 The reason they believe that camouflage plays a role is that the peacock train (i.e. undeployed tail feathers) is mostly green and supposedly provides camouflage when it hides in trees. However, the theory of camouflage also has serious problems. Firstly, the tail makes the peacock more conspicuous on the ground, which is arguably where the greatest danger is to be found. Secondly, camouflage does not explain how the subtle eye patterns could have evolved. Thirdly, if the function of camouflage were really effective then the peahen should also have such a tail.
(Iii) Recognition
Some evolutionists believe that the colour and pattern of the peacock tail has the sole function of making the peacock recognizable to the peahen.25 However, this theory cannot begin to explain the origin of the subtle aesthetic features of the peacock.
Added beauty
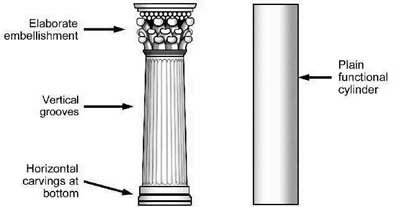
Figure 8. Added beauty in a column.
The beauty of the peacock tail can be termed ‘added beauty’ because it appears to be surplus to that necessary to survive. In other words, the beauty of the peacock tail is not a by-product of the function of the tail. Added beauty can be a powerful evidence of design because it is a common hallmark of an intelligent designer. The hallmark of added beauty can be seen in all kinds of human design. For example, an architect often adds decorative features to the different parts of a building. The adding of beauty to a column is illustrated in Figure 8 (left), which compares a classical column with a plain functional cylinder. The decorative features of the classical column have the sole function of providing a beautiful spectacle. But they also present evidence that an intelligent designer has designed the column. So also the added beauty of a peacock tail reveals an intelligent designer.
Most evolutionists accept that creatures like the peacock have added beauty. This is why the peacock tail feathers are referred to as decorative feathers in standard biology textbooks. Darwin said this about beauty in nature: ‘A great number of male animals … have been rendered beautiful for beauty’s sake’; ‘the most refined beauty may serve as a charm for the female, and for no other purpose.’26 Considering that evolutionists recognize added beauty in nature, and considering that added beauty is very much a hallmark of an intelligent designer, beauty in nature must be seen as an important evidence of design. A study of added beauty in nature has been described in my book Hallmarks of Design.27
Conclusion
There are many beautiful features in the peacock tail such as bright iridescent colours, intricate patterns and the fan-formation of the displayed feathers. The mechanisms that are responsible for producing these beautiful features are very sophisticated. In particular, the barbules contain an astounding level of precision design in order to produce optimum thin-film interference.
There are several serious problems with the evolutionary theory of sexual selection. There is no satisfactory explanation of how the sexual selection cycle can start or why the peahen should prefer beautiful features. In addition, there is irreducible complexity in both the physical structure of the feather and in the beautiful patterns.
Darwin once said, ‘The sight of a feather in a peacock’s tail, whenever I gaze at it, makes me feel sick!’28 If Darwin knew about the modern discoveries re the complexities of the peacock tail, he would have even greater reason to feel sick. In contrast, when Christians look at the tail of the peacock they can feel a wonderful assurance that there is a Creator who is infinite in wisdom and that the Creator has been very good to mankind in putting such beauty into the world.
Acknowledgements
I would like to thank Des O’Neil from Bristol University for taking pictures of the barbules under the microscope.
Footnotes
- Some of the peacock display feathers have a few centimetres of fluffy barbs at their root to perform the function of thermal insulation. However, the fluffy barbs are such a minor detail that the description of ‘display feather’ is generally used by biologists.
- King, A.S. and McLelland, J., Birds—Their Structure and Form, Bailliere Tindall, p. 15, 1984.
- Sager, E., Morphologishce Analyse der Musterbildung beim Pfauenrad, Revue Suisse de Zoologie, Geneve 62(2):116, 1955.
- Mason, C.W., Structural colours in feathers II, J. Physical Chemistry 27:440, 1923.
- Mason, p. 416.
- Mason, p. 416.
- Mason, p. 417.
- Mason, p. 443.
- Mason, p. 442.
- Mason, p. 442.
- Marshall, A.J. (Ed.), Biology and Comparative Physiology of Birds, Academic Press, London, p. 225, 1960.
- Darwin, C. The Descent of Man, John Murray, London, p. 412, 1871.
- Cronin, H., The ant and the peacock, Cambridge University Press, Cambridge, Great Britain, p. 118, 1991.
- Darwin, C., The Descent of Man, John Murray, London, p. 349, 1888.
- Cronin, p. 202.
- Andersson, M, Sexual Selection, Princeton University Press, Princeton, p. 336, 1994.
- Cronin, p. 222.
- Cronin, p. 202.
- Cronin, p. 186.
- <www.talkorigins.org/faqs/faq-intro-to-biology.html>, p. 7, 2001.
- Darwin, C., p. 412.
- Mason, p. 444.
- Cronin, pp. 221–223.
- Clarke, D. (Ed.), Wildlife Fact File, International Publishers Ltd, London, 1972.
- Dawkins, R., The Blind Watchmaker, Penguin, London, p. 199, 1986.
- Quoted from: Cronin, p. 183.
- Burgess, S.C., Hallmarks of Design, Day One Publications, Epson, Surrey, 2000.
- Darwin, F., (Ed), Letter to Asa Gray, dated 3 April 1860, The Life and Letters of Charles Darwin, John Murray, London, Vol. 2, p. 296, 1887; 1911 Edition, D. Appleton and Company, New York and London, Vol. 2, pp. 90–91.
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis